A Simple and Efficient Method to Detect Nuclear Factor Activation in Human Neutrophils by Flow Cytometry
Summary
Neutrophils are the most abundant leukocytes in blood. Neutrophils possess transcriptionally regulated functions such as production of proinflammatory cytokines and inhibition of apoptosis. These functions can be studied with the method presented here, which allows detection and quantification of nuclear factors by flow cytometry in isolated nuclei
Abstract
Neutrophils are the most abundant leukocytes in peripheral blood. These cells are the first to appear at sites of inflammation and infection, thus becoming the first line of defense against invading microorganisms. Neutrophils possess important antimicrobial functions such as phagocytosis, release of lytic enzymes, and production of reactive oxygen species. In addition to these important defense functions, neutrophils perform other tasks in response to infection such as production of proinflammatory cytokines and inhibition of apoptosis. Cytokines recruit other leukocytes that help clear the infection, and inhibition of apoptosis allows the neutrophil to live longer at the site of infection. These functions are regulated at the level of transcription. However, because neutrophils are short-lived cells, the study of transcriptionally regulated responses in these cells cannot be performed with conventional reporter gene methods since there are no efficient techniques for neutrophil transfection. Here, we present a simple and efficient method that allows detection and quantification of nuclear factors in isolated and immunolabeled nuclei by flow cytometry. We describe techniques to isolate pure neutrophils from human peripheral blood, stimulate these cells with anti-receptor antibodies, isolate and immunolabel nuclei, and analyze nuclei by flow cytometry. The method has been successfully used to detect NF-κB and Elk-1 nuclear factors in nuclei from neutrophils and other cell types. Thus, this method represents an option for analyzing activation of transcription factors in isolated nuclei from a variety of cell types.
Introduction
Neutrophils are the most abundant leukocytes in peripheral blood 1. During inflammation and infection neutrophils are the first cells to appear at the affected site where they act as the first line of defense 2. Neutrophils possess several antimicrobial mechanisms 3 including phagocytosis, production of reactive oxygen species, release of lytic enzymes by degranulation, and production of proinflammatory cytokines 4,5. Neutrophils are short-lived cells that get rapidly activated through signaling from various cell surface receptors. Although neutrophils have been considered terminal cells due to their short life and because they undergo apoptosis unless activated during the inflammatory process 6, it is now clear that they can also modify their phenotype by changing the level of transcription of particular genes. The production of cytokines 5 and the inhibition of apoptosis 7,8 are two important activation-dependent cell functions regulated at the level of transcription in neutrophils. Nuclear factor κB (NF-κB) participates in the transcriptional control of cytokine production 4 and in the regulation of cell survival and apoptosis 9-11 in various cell types.
The signaling pathways that lead to nuclear factor activation are usually studied by reporter gene assays or by electrophoretic mobility shift assays (EMSA). However, because neutrophils are short-lived cells, the study of transcriptionally regulated responses in these cells cannot be performed with reporter gene assays, since there are no efficient techniques for neutrophil transfection. EMSA assays have been used in neutrophils to explore nuclear factor activation 12,13; however, this methodology is complicated and expensive because it involves the use of radioactive material. Nucleofection is another technique that has been used successfully to transfect monocytes 14. Thus, at least in theory, nuclear factor activation could be detected in neutrophils by transfection (despite low efficiency). However, this technique would be more expensive, time-consuming and probably less quantitative. Microscopic analysis of immunostained cells could also be used to detect nuclear factors in the nucleus. Indeed, we have detected NF-κB translocation into the nucleus this way 15. Unfortunately, this technique is also time-consuming, less quantitative, and subject to the observer’s bias.
Here, we present a simple and efficient method that allows detection and quantification of nuclear factors in isolated and immunolabeled nuclei by flow cytometry. We describe techniques to isolate neutrophils from human peripheral blood, stimulate these cells via integrins or Fc receptors with anti-receptor antibodies, isolate and immunolabel nuclei, and analyze nuclei by flow cytometry (Figure 1). The method has been successfully used to detect NF-κB 15 and Elk-1 16 nuclear factors in neutrophil nuclei. The sensitivity of this method allows detection of small changes in nuclear factor levels in the nucleus. This method can also be used to analyze the level of transcription factors in nuclei from other cell types.
Protocol
1. Isolation of Neutrophils (PMN) from Human Blood
- Use about 20 ml human blood with heparin (10 U/ml) as anticoagulant. Blood was collected from adult healthy volunteers by venopuncture. All experiments were done under approval of the Bioethics Committee at the Instituto de Investigaciones Biomédicas – UNAM.
- Put 2 ml of 6% dextran T500 in PBS into a 15 ml conical centrifuge tube and add 10 ml of blood. Mix by inverting the tube two or three times and let it sit for 45 min to allow for erythrocyte sedimentation.
- In a fresh 15 ml conical centrifuge tube put 5 ml Ficoll-Paque.
- Take the leukocyte-rich plasma that formed above sedimented erythrocytes, and carefully pipette it on top of the Ficoll-Paque forming a second layer. Make sure there are two phases.
- Centrifuge at 516 x g for 20 min at 4 °C. After centrifugation, at the interphase of plasma and Ficoll-Paque there is a layer of mononuclear cells. Neutrophils are at the bottom of the tube.
- Eliminate the supernatant, break the cell pellet by tapping the tube against a rack, and resuspend the cells by adding 10 ml of cold (4 °C) PBS. Note: Resuspending the cells by pipetting up and down is not recommended since this is too damaging to the cells.
- Transfer the cell suspension to a 50 ml conical centrifuge tube and centrifuge at 290 x g for 5 min at 4 °C.
- Break the cell pellet as before and add 10 ml of cold (4 °C) hypotonic solution (0.2% NaCl, 1% BSA, 20 mM Hepes, pH = 7.4) to lyse erythrocytes. Mix by swirling the tube gently by hand for exactly one minute.
- Add 10 ml of cold (4 °C) hypertonic solution (1.6% NaCl, 1% BSA, 20 mM HEPES, pH = 7.4), mix and count the cells using a hemocytometer (usually >95% are PMN).
Note: Keep the tube with the cell suspension on ice while counting the cells. - Centrifuge as before and resuspend PMN at 107 cell/ml in cold (4 °C) PBS. Keep on ice.
Note: Neutrophils remain viable and functional at 4 °C for about 6 to 8 hr.
2. Activating PMN
- Put 100 μl of PMN suspension (1 x 107cell/ml) into a 1.5 ml Eppendorf tube.
Note: Sample tubes should be done at least in duplicates. Usual negative controls should include first antibody only, and secondary antibody only. - Add the corresponding anti-integrin or anti-Fc receptor monoclonal antibody (mAb) at 10 μg/ml and incubate on ice for 15 min.
- Wash PMN by adding 1 ml of cold (4 °C) PBS, centrifuging at 1,743 x g (4,500 rpm) in a microcentrifuge, and aspirating the supernatant.
- Break the cell pellet by tapping the bottom of the tube against a rack, add 1 ml cold PBS, and wash two more times as in previous step.
Note: These washes remove unbound antibody. - Resuspend PMN in same (initial) volume of warm (37 °C) PBS containing 60 μg/ml of F(ab’)2 goat anti-mouse IgG.
Note: The secondary anti-mouse IgG antibody will bind to the first anti-receptor antibody and will cause crosslinking of the receptor. - Incubate at 37 °C for 1 to 20 min, depending on the nuclear factor of interest. For NF-κB usually 15 min, and for Elk-1 usually 5 min.
Note: Incubating the cells at 37 °C induces receptor crosslinking, which cannot take place at 4 °C. - Add 1 ml cold PBS and centrifuge 3 min at 1,743 x g in microcentrifuge.
- Remove supernatant
- Freeze PMN immediately in dry-ice/ethanol bath and keep them there for 10 min.
3. Isolation and Fixation of Nuclei
- Resuspend frozen PMN pellet in 100 μl cold (4 °C) hypotonic buffer (10 mM HEPES, 10 mM KCl, 1.5 mM MgCl2, and 1 mM freshly-added DL-dithiothreitol [DTT]; pH = 7.9). It is recommended to take the tubes out of the dry-ice/ethanol bath one by one, and to wipe clean the bottom of the tube before adding the hypotonic buffer.
- Place on ice, and check for nuclei integrity by staining an aliquot of the nuclei suspension with trypan blue. Nuclei look round and blue. Intact PMN do not get stained, and cell debris appears as blue particles of irregular shape.
Note: If nuclei suspension contains many intact cells or debris, it is better to discard the preparation and repeat the procedure with another sample. - Centrifuge nuclei at 775 x g (3,000 rpm) in microcentrifuge for 10 min inside a cold room. At this point nuclei are very fragile and extra care should be taken in keeping the samples cold and not centrifuging at excessive forces.
- Remove supernatant by very careful pipetting.
- Add 100 μl of cold 4% paraformaldehyde in PBS to fix nuclei.
Note: The pellet is very loose and adding the buffer is enough to resuspend the nuclei. Pipetting up and down should be avoided. - Incubate on ice for 20 min.
- Immunolabel nuclei for flow cytometry or keep them for up to 24 hr at 4 °C.
4. Nuclei Immunolabeling for Flow Cytometry Analysis
- Centrifuge nuclei at 1,743 x g (4,500 rpm) in microcentrifuge for 1 min, and remove supernatant by very gentle pipetting.
- Add 100 μl of cold (4 °C) 0.1% Triton X-100, 4% paraformaldehyde in PBS to permeabilize nuclei. Incubate in ice for 10 min.
- Centrifuge permeabilized nuclei at 1,743 x g in microcentrifuge and carefully remove the supernatant. At this point nuclei pellets do not attach well at the bottom of the tube. It is recommended to centrifuge again if the pellet gets loose.
- Resuspend nuclei in 500 μl of cold 4% fetal bovine serum (FBS) in PBS to block nonspecific binding sites, and incubate on ice for 20 min. Centrifuge nuclei at 1,743 x g for 3 min in microcentrifuge and carefully remove the supernatant.
- Resuspend nuclei in 100 μl of cold PBS containing 4% FBS and 2.5 μg/ml of the mAb against the nuclear factor of interest. Incubate on ice for 20 min.
Note: Usual negative controls should include anti-nuclear factor antibody only, and FITC-labeled secondary antibody only. - Wash twice with 500 μl of cold PBS with 4% FBS and centrifuging at 1743 x g for 3 min.
- Remove the supernatant very carefully, resuspend nuclei in 100 μl of cold PBS containing 4% FBS and 10 μg/ml of the corresponding FITC-labeled secondary antibody, and incubate on ice for 20 min.
- Wash nuclei twice with 500 μl of cold PBS with 4% FBS as in previous step.
- Resuspend nuclei in 400 μl of cold 4% paraformaldehyde in PBS.
Note: Nuclei are placed in paraformaldehyde to allow the sample to be stored for several days without contamination problems that can occur in the presence of serum. - Analyze immediately by flow cytometry or store in the dark at 4 °C for up to three days.
5. Flow Cytometry Analysis
- Analyze immunolabeled nuclei in a flow cytometer such a FACScan (Becton Dickinson; Franklin Lakes, NJ) or similar apparatus.
- Adjust acquisition settings to: FSC at log-scale at 10-1, SSC at log-scale at 196, and gate nuclei in a dot-plot.
- Acquire ten thousand nuclei per sample.
- Analyze fluorescence of FITC-stained nuclei through the FL-1 channel (506 nm) set at log-scale at 400.
Representative Results
The purification method described here usually provides unstimulated neutrophils (PMN) with purity greater than 95% (Figure 1A). Isolated PMN can then be stimulated by crosslinking particular receptors with specific monoclonal antibodies. We have stimulated PMN through Fc receptors and integrins (Figure 1B). Once stimulated, PMN are lysed and nuclei are isolated with high yields. Nuclei are then immunolabeled for a particular nuclear factor, such as the nuclear factor κB (NF-κB), with specific antibodies (Figure 1B).
Nuclei can be easily recognized as a different population from intact PMN in the flow cytometer with a dot-plot (Figure 2A). In resting PMN a basal level of NF-κB is found in the cell nucleus (Figure 2B). Upon stimulation by crosslinking β1 integrins, NF-κB is translocated to the nucleus and this increment in nuclear NF-κB is detected as an increase in fluorescence (Figure 2B). Similarly, stimulation of PMN by crosslinking β2 integrins also induced NF-κB activation as indicated by an increase in fluorescence (Figure 2C). The sensitivity of this method allows detection of small changes in nuclear factor levels as evidenced by the fact that β1 integrins, which bind extracellular matrix proteins such as fibronectin, induced stronger NF-κB activation than β2 integrins, which bind to adhesion molecules on other cells (compare Figures 2B and 2C).
This method has also been successfully used to detect NF-κB 15 and Elk-1 16 nuclear factors in isolated nuclei after Fc receptor stimulation of PMN. This method is simple and economical, thus it can easily be modified to analyze nuclear factors in other cell types.
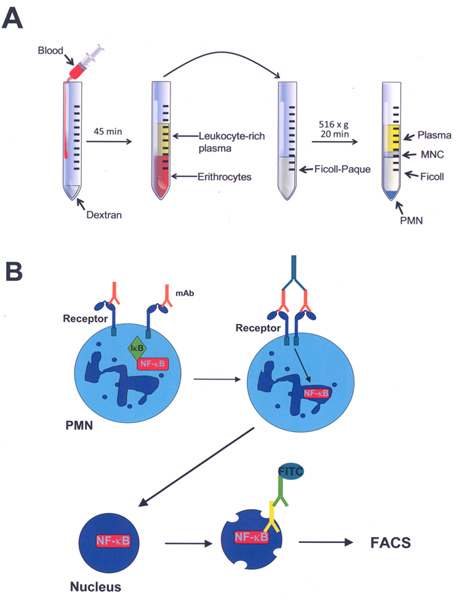
Figure 1. Schematic representation of neutrophil (PMN) purification from blood, stimulation of cells, isolation of nuclei, and immunolabeling of nuclear factors in isolated nuclei. A) Heparin-treated blood is mixed with dextran to agglutinate erythrocytes. After erythrocytes sediment, the leukocyte-rich plasma is on top of the red cells. This leukocyte-rich plasma is transferred and layered on top of Ficoll-Paque in another tube. After centrifugation, a layer of mononuclear cells (MNC) is found at the interphase of plasma and Ficoll-Paque. PMN are found at the bottom of the tube. B) Isolated PMN are stimulated by crosslinking cell-membrane receptors with specific monoclonal antibodies (mAb) (orange) and a secondary antibody (dark green). Nuclear factor kappa B (NF-κB) separates from its inhibitor IκB and translocates into the nucleus. Nuclei are then isolated, permeabilized and immunolabeled against NF-κB or another nuclear factor with a specific mAb (yellow), and a FITC-labeled secondary antibody (light green). Fixed nuclei are analyzed by flow cytometry (FACS). Click here to view larger figure.
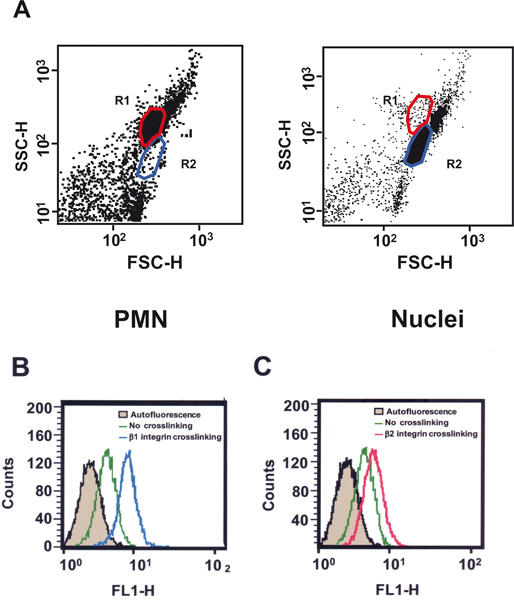
Figure 2. Analysis of NF-κB activation in neutrophils (PMN). A) Intact PMN (left panel) or isolated nuclei (right panel) appear as two distinct populations in flow cytometry dot-plot graphs. Cells and nuclei can be analyzed independently by creating different gates, R1 (red) for intact cells, and R2 (blue) for isolated nuclei. B and C) Nuclei isolated from PMN were immunolabeled for NF-κB (p50 subunit), before (green dashed line), or after cells were activated by cross-linking β1 integrins (B) with the specific monoclonal antibody TS2/16 (blue line); or β2 integrins (C) with the specific monoclonal antibody IB4 (red line). The gray area is the basal fluorescence from the FITC-labeled secondary antibody alone.
Discussion
The purification method described here allows the isolation of unstimulated neutrophils (PMN) with purity greater than 95% (assessed by microscopic observation), in a short time. Sometimes neutrophils can be contaminated by erythrocytes if the latter are not lysed completely. This does not usually affect the technique, since erythrocytes and PMN can easily be distinguished as distinct cell populations by flow cytometry. Isolated PMN can then be stimulated by crosslinking particular receptors with specific monoclonal antibodies. In fact, PMN can also be stimulated by pharmacological means or by adhesion to various substrates or other cells. The method presented here can be used to analyze signaling pathways that lead to changes of nuclear factors in PMN nuclei after cells are activated by practically any relevant biological stimulus.
Once PMN are stimulated, the method described in this paper allows for the rapid and efficient purification of nuclei and for the detection, in these nuclei of NF-κB by immunofluorescence and flow cytometry analysis. After PMN stimulation of certain receptors, NF-κB in the cytoplasm is released from its inhibitor IκB, and then traslocated into the nucleus (Figure 1B). Thus, the level of NF-κB activation is determined by the amount of NF-κB present in the nucleus. Because this method can detect and measure the amount of NF-κB in the nucleus, it can easily determine activation of this nuclear factor. The method can in a similar manner detect activation of other nuclear factors, when their activation is linked to changes in nuclear levels. Activating changes of nuclear factors can be due to changes in the amount of factor present in the nucleus, as it is the case for NF-kB; or changes such as phosphorylation, in the molecular structure of the factor, as it is the case for Elk-1. This method can be used to detect changes of nuclear levels of any molecule for which a specific antibody is available.
This method yields a pure nuclei population in a simple and fast manner. Cells are frozen and lysed in a hypotonic buffer that burst the cells open and leaves intact nuclei. Intact PMN or isolated nuclei usually appear as two distinct populations in flow cytometry dot-plot graphs. Cells and nuclei can be analyzed independently by creating different gates for intact cells, and for isolated nuclei. In an intact cell sample a gate is created around the area where most cells appear. Similarly, with a nuclei sample a gate is created around the area where most nuclei appear. Normally these two gates are in different places of the dot-plot graph. Particles that are sometimes detected out of these gates, are usually cell or nuclei debris, and they are left out of the analysis. If the cell and nuclei gates overlap, it usually means that the nuclei preparation is not clean. Analysis of this nuclei preparation is not reliable, and it is better to repeat the procedure with freshly made buffers.
The method was developed to work with human PMN, but it has also been successfully applied to various cell lines of leukocyte origin 15. The method can easily be adapted to isolate nuclei from various cell types. It is important to begin the procedure with a minimum of 1 x 106 cells, to obtain enough nuclei for an efficient flow cytometry analysis. If small numbers of nuclei are obtained at the end, it is better to routinely start the method with 3 x 106 cells.
During the lysis step, frozen cells should not be allowed to thaw. This results in low yield due to cell degradation. Thus, it is recommended to take one tube at a time out of the dry-ice/ethanol bath to lyse cells. Also, it is important to wipe clean the outside of the tube before adding the hypotonic lysis buffer. When this is not done, the buffer sometimes freezes in the tube, giving an inefficient cell lysis and resulting in nuclei preparations that are contaminated by intact cells. Still, this is not a serious problem given that in most cases intact cells and nuclei can be separated by flow cytometry.
Because nuclei are very fragile before they get fixed, it is important to have all buffer solutions cold at 4 °C by keeping them in ice. After centrifugation, nuclei pellets are sometimes loose at the bottom of the tube, so extra care should be taken when removing the supernatant. This is better done with a micropipette than with vacuum aspiration. On occasion, the tube can be centrifuged again, but the recommended centrifugation forces should not be excessive because this damages the nuclei.
The method detected NF-κB activation using antibodies specific to the p50 subunit (Figure 2), and also the p65 subunit 15 of this nuclear factor. Also, quantification of receptor-initiated nuclear activation of ERK and Elk-1 was also successfully achieved through this method 16, using the corresponding specific antibodies against these molecules. In addition, the sensitivity of this method allowed detection of small changes in NF-κB nuclear levels, due to differential stimulation of PMN by different types of integrins (Figure 2), and also due to pharmacological inhibition of Fc receptor-initiated signaling pathways 16. The combination of a good specific antibody and an efficient fluorescently-labeled secondary antibody provides great sensitivity and allows for the detection of small changes in the amount of the nuclear factor of interest inside nuclei. This permits a quantitative comparison of the levels of the nuclear factor before and after different stimulation conditions. Also, this method can detect in isolated nuclei, virtually any molecule against which a good specific antibody is available. Furthermore, the immunolabeling part can be simplified by using an antibody directly labeled with the fluorochome, which in addition will improve sensibility. Finally, the method presents great flexibility because many different fluorochomes can be used. Thus, this method can be applied to many different signal transduction studies that involve changes of protein levels in the nucleus.
In conclusion, the broad applicability and sensitivity of the method described in this paper, place it as a simple and economical option for signal transduction studies that involve changes of protein levels in the nucleus of a variety of cell types.
Disclosures
The authors have nothing to disclose.
Acknowledgements
The authors would like to thank Nancy Mora for her technical assistance.
This work was funded by research grants 48573-M and 168098 from Consejo Nacional de Ciencia y Tecnologia, Mexico, and by grants IN212308 and IN205311-2 from Direccion General de Asuntos del Personal Academico, Universidad Nacional Autonoma de Mexico, Mexico.
Materials
| REAGENTS | |||
| Heparin | PiSA (Mexico) | ||
| Dextran T500 | Pharmacosmos A/S (Holbaek, Denmark) | T1-Dextran T500 | |
| Ficoll-Paque | Pharmacia | 17-0320-01 | |
| Sodium chloride | Sigma | S7653 | |
| Sodium phosphate monobasic | Sigma | S9638 | |
| Sodium phosphate dibasic | Sigma | S9390 | |
| Bovine serum albumin (BSA) | Sigma | A2153 | Cohn Fraction V |
| HEPES | Sigma | H3375 | |
| Potassium chloride | Sigma | P9541 | |
| Magnesium chloride anhydrous | Sigma | M8266 | |
| DL-dithiothreitol (DTT) | Sigma | D9163 | |
| Trypan Blue (0.4 % solution) | Sigma | T8154 | |
| Paraformaldehyde | Sigma | P6148 | |
| Triton X-100 | Sigma | X100 | |
| Fetal bovine serum (FBS) | GIBCO | 10437-028 | |
| Monoclonal antibody IV.3 | Medarex (Annandale, NJ) | 025-1 | Human-specific anti-FcRII (CD32) |
| Monoclonal antibody 3G8 | Medarex (Annandale, NJ) | 028-2 | Human-specific anti-FcRIII (CD16) |
| Monoclonal antibody TS2/16 | Dana Farber Cancer Research Institute (Boston, MA) | Donated by Dr. Martin Hemler | Human-specific anti-β1 integrin (CD29) |
| Monoclonal antibody IB4 | University of California, San Francisco | Donated by Dr. Eric J. Brown | Human-specific anti-β2 integrin (CD18) |
| F(ab’)2 goat anti-mouse IgG | Cappel (Aurora, OH) | 55468 | |
| FITC-conjugated F(ab’)2 goat anti-mouse IgG | Cappel (Aurora, OH) | 55522 | |
| FITC-conjugated F(ab’)2 goat anti-rabbit IgG | Cappel (Aurora, OH) | 55665 | |
| Anti-NF-κB p50 | Santa Cruz Biotechnology (Santa Cruz, CA) | sc-114 | Rabbit polyclonal antibody |
| Anti-NF-κB p65 | Santa Cruz Biotechnology (Santa Cruz, CA) | sc-109 | Rabbit polyclonal antibody |
| EQUIPMENT | |||
| 15-ml centrifuge tube | Corning | 430791 | |
| 50-ml centrifuge tube | Corning | 430291 | |
| Centrifuge, Sorvall Tabletop | Dupont Instruments | RT 6000D | |
| pH-meter | Corning | 340 | |
| Pipetman pipette P-20 | Gilson | F123600 | |
| Pipetman pipette P-200 | Gilson | F123601 | |
| Pipetman pipette P-1000 | Gilson | F123602 | |
| Hemocytometer | Fisher Scientific | 0267110 | |
| Microscope | Nikon | Eclipse E600 | |
| Inverted microscope | Nikon | TMS | |
| Water Bath Incubator | Fisher Scientific | 2IS-M | |
| Microcentrifuge | Eppendorf | 5414C | |
| Microcentrifuge | Eppendorf | 5418 | |
| Flow Cytometer | Becton Dickinson (Franklin Lakes, NJ) | FACScalibur |
References
- Sendo, F., et al. Regulation of neutrophil apoptosis: its biological significance in inflammation and the immune response. Human Cell. 9, 215-222 (1996).
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity. 33, 657-670 (2010).
- Naussef, W. M. How human neutrophils kill and degrade microbes. An integrated view. Immunol. Rev. 219, 88-102 (2007).
- Scapini, P., et al. The neutrophil as a cellular source of chemokines. Immunol. Rev. 177, 195-203 (2000).
- Hamilton, T., et al. Cell type- and stimulus-specific mechanisms for post-transcriptional control of neutrophil chemokine gene expression. J. Leukoc. Biol. 91, 377-383 (2012).
- Simon, H. U. Neutrophil apoptosis pathways and their modifications in inflammation. Immunol. Rev. 193, 101-110 (2003).
- Akgul, C., Moulding, D. A., Edwards, S. W. Molecular control of neutrophil apoptosis. FEBS Lett. 487, 318-322 (2001).
- Witko-Sarsat, V., Pederzoli-Ribeil, M., Hirsch, E., Sozzani, S., Cassatella, M. A. Regulating neutrophil apoptosis: new players enter the game. Trends Immunol. 32, 117-124 (2011).
- Green, D. R. Death and NF-κB in T cell activation: life at the edge. Mol. Cell. 11, 551-552 (2003).
- Papa, S., Zazzeroni, F., Pham, C. G., Bubici, C., Franzoso, G. Linking JNK signaling to NF-κB: a key to survival. J. Cell Sci. 117, 5197-5208 (2004).
- Valente, P., et al. TNF increases camptothecin-induced apoptosis by inhibition of NF-κB. Eur. J. Cancer. 39, 1468-1477 (2003).
- Choi, M., et al. Inhibition of NF-κB by a TAT-NEMO-binding domain peptide accelerates constitutive apoptosis and abrogates LPS-delayed neutrophil apoptosis. Blood. 102, 2259-2267 (2003).
- Wang, K., et al. Inhibition of neutrophil apoptosis by type 1 IFN depends on cross-talk between phosphoinositol 3-kinase, protein kinase C-d, and NF-κB signaling pathways. J. Immunol. 171, 1035-1041 (2003).
- Schnoor, M., et al. Efficient non-viral transfection of THP-1 cells. J. Immunol. Meth. 344, 109-115 (2009).
- Garcia-Garcia, E., Rosales, C. Nuclear factor activation by FcγR in human peripheral blood neutrophils detected by a novel flow cytometry-based method. J. Immunol. Meth. 320, 104-118 (2007).
- Garcia-Garcia, E., Nieto-Castaneda, G., Ruiz-Saldana, M., Mora, N., Rosales, C. FcγRIIA and FcγRIIIB mediate nuclear factor activation through separate signaling pathways in human neuthophils. J. Immunol. 182, 4547-4556 (2009).
