Results shown below represent separate experiments in which either airway or vascular compartments were seeded with rhesus macaque bone marrow-derived mesenchymal stem cells or rhesus lung-derived microvascular endothelial cells, respectively, that were isolated and characterized as previously described16-18.
Throughout the decellularization process, macaque lungs displayed a progressive whitening culminating in a translucent appearance at the end of the process; however, the lungs maintained their gross anatomical features and remain largely elastic and able to produce natural recoil after inflation with liquid (Figures 3A and 3B). At the microscopic level, the histologic ultrastructure remained intact after decellularization; that is, bronchioles, respiratory bronchioles, alveolar sacs, blood vessels, and capillaries were still distinguishable quite clearly by low-power microscopy (Figures 3C and 3D). Histological microanatomy, however, demonstrated that while the gross anatomy and ultrastructure of the lung were mildly disturbed by decellularization, the tissue completely lacked intact cells (Figures 3E and 3F).
Cellular DNA that was released as cells were lysed by the detergents was degraded by DNase treatment and efficiently removed by subsequent washing steps. DNA was extracted from native and decellularized macaque lung tissues using Qiagen's DNeasy kit. As seen in Figure 4A, only trace amounts of DNA remained in decellularized tissues; moreover, the trace DNA (concentrated by alcohol precipitation and visualized in a 0.8% agarose gel) was composed of mostly low molecular weight degraded fragments (Figure 4A, inset). The efficiency of cellular protein removal was assessed by Western blot analyses of both native and decellularized lung protein lysates using an antibody to β-actin. Briefly, equal protein amounts (25 µg) in triplicate for each native and decellularized sample were subjected to PAGE at 200 V for 1 hr. Proteins were transferred to nitrocellulose using an Invitrogen iBlotter. Blots were probed with a mouse anti-human β-actin primary antibody (IgG1) followed by a goat anti-mouse IgG-HRP secondary antibody. Equal protein loading was confirmed by staining an identical membrane with Ponceau-S and performing densitometry to determine total protein stain intensity between native and decellularized samples (not shown). The blot was developed using ECL chemiluminescent detection reagent (Invitrogen) and imaged using the ImageQuant LAS 4000 system (GE). β-actin was easily detected in native lung lysates but not in decellularized lung lysates, suggesting that decellularization dramatically depleted cells and removed cell-associated protein material (Figures 4B).
An example of the assembled bioreactor with organs installed is shown in Figure 5. A cell culture water pan easily held all three chambers and provided containment in the event that a leak occurred. The latex tubing used as sealing gaskets for the lids of the bioreactor chambers also functioned to prevent kinks in the tubing by providing added rigidity in areas where the tubing would otherwise fold on itself. The bioreactor, 6-port syringe pump, and peristaltic pump fit easily into a standard cell culture incubator. The electronic components were powered via a flat-cord electrical extension that is sandwiched between the inner incubator door and the rubber seal. The incubator was maintained at 37°C, 5% CO2 throughout experiments.
Fourteen days after airway seeding with macaque BMSC (1.0 x 106 cells/ml) and bioreactor culture with ~1 inspiration/expiration cycle every 2 min, the parenchyma of decellularized macaque lungs was effectively recellularized (Figure 6A). BMSCs lined the alveolar septae while maintaining a clear and open alveolar lumen. Inoculation of cells into the trachea (or bronchioles for single lobes) was an efficient route for inoculating distal lung tissues as is evident in by the recellularization of alveoli near the pleura at the extremities of the lung. The denuded matrix of large airways was also recellularized by BMSC using the bioreactor. Figure 6B shows the luminal surface of a mainstem bronchus that was lined with a monolayer of squamous-like BMSC after 14 days of culture in the bioreactor.
Histological analysis of a decellularized rhesus lung lobe five days after seeding the vasculature with microvascular endothelial cells (8.0 x 104 cells/ml) and providing constant vascular perfusion with endothelial culture medium at 5 ml/min, revealed cells lining the small vasculature in the lung parenchyma (Figures 6C and 6D). In some instances, cells appeared to be attached to the matrix via several cellular projections across the lumen (Figure 6C) while other cross sections of vessels showed cells lining the endothelial surface with clear lumen (Figure 6D).
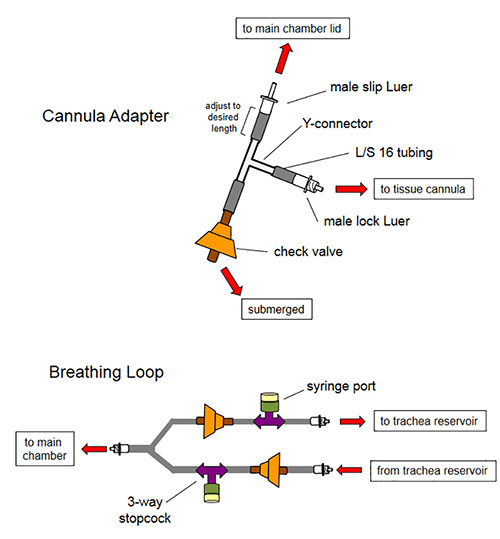
Figure 1. Assembly schematic for tracheal and vascular cannulae adapters and breathing loop. Cannulae adapters allow removal of air from the cannula tubing after the tissues are installed in the bioreactor. A one-way check valve allows medium to be aspirated from the main chamber as to not produce undue force on the tissue airway or vasculature. Liquid traveling toward the tissue will be diverted into the airway or vasculature because reverse pressure on the check valve will cause it to close. This same principle produces the main function of the breathing loop; here, air is removed by aspirating liquid through the one-way valves using a needle and syringe via the in-line injection ports.
Click here to view larger image.
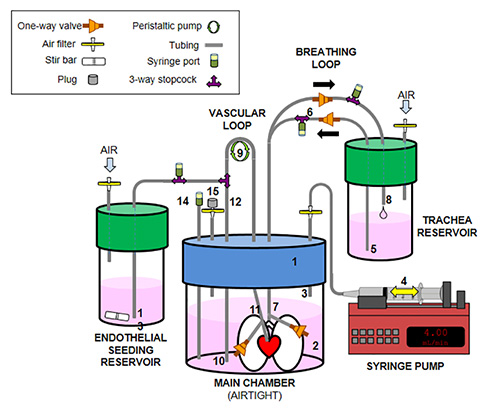
Figure 2. Assembly schematic for the large-organ bioreactor. The following components constitute the main functionality of the lung bioreactor: (1) Modified 2 L jar lid; (2) main chamber containing culture media (3) air intake/outlet for negative pressure ventilation; (4) syringe pump cycles air in and out of main chamber; (5) culture media moves from trachea reservoir into the lung airway when syringe withdraws air from main chamber; (6) one-way valves allow constant cycling of culture media into and out of lung via a breathing loop — airway cells are directly injected into the breathing loop to inflate the lung statically for 18 hr before mechanical cyclic ventilation; (7) media enters lung via the trachea and lung inflates; (8) when syringe pump returns air to the main chamber, lungs deflate and return culture media to the trachea reservoir; (9) a peristaltic pump moves culture media from the main chamber (10) into the pulmonary artery (11), through the lung vasculature, and back into the main chamber; (12) to seed the vasculature with endothelial cells, the endothelial seeding reservoir is temporarily connected to the vascular loop via a 3-way stopcock, the position of the valve is moved so that media containing endothelial cells in the endothelial seeding reservoir is drawn up (13) through the peristaltic pump into the pulmonary artery (11) and lung vasculature. When endothelial seeding is complete (volume in the endothelial seeding reservoir is completely transferred to the main chamber through the lung vasculature), the stopcock is returned to its original position to allow media to be drawn up from the main chamber (10). (14) Injection port for changing main chamber media; (15) air filter is capped during operation; the cap is removed to allow changing of air inside the main chamber.
Click here to view larger image.
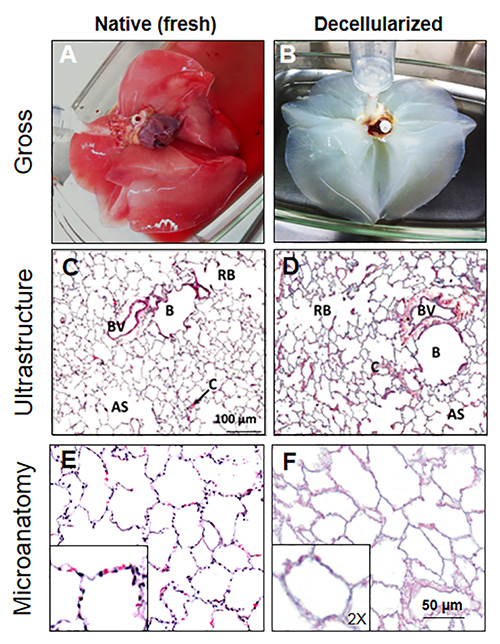
Figure 3. Decellularization of rhesus macaque lungs. A) Native lungs obtained from necropsy are perfused via the pulmonary artery to remove excess blood from the pulmonary capillary bed. B) After treatment with Triton X-100, Deoxycholic acid, hypertonic NaCl, and DNase, the lungs become white and translucent, indicative of decellularization. C) The ultrastructure of the native lung is represented in normal histology (H&E) by the appearance of bronchioles "B", blood vessels "BV", respiratory bronchioles "RB", capillaries "C", and alveolar sacs "AS". D) Lung ultrastructure is largely unaffected by decellularization, as all structural components of normal lung histology are visible after decellularization. E) H&E stain of normal native lung parenchyma shows the lacy appearance of alveoli in a 5 µm paraffin section with alveolar epithelial cells and some remaining red blood cells. F) The parenchyma of decellularized lungs contains alveoli that are intact relative to its native counterpart; however, these alveoli are completely devoid of cells as noted by the absence of dark-staining nuclei or red blood cells. Insets represent 2X magnification relative to their respective panels. Reprinted with permission from reference19.
Click here to view larger image.
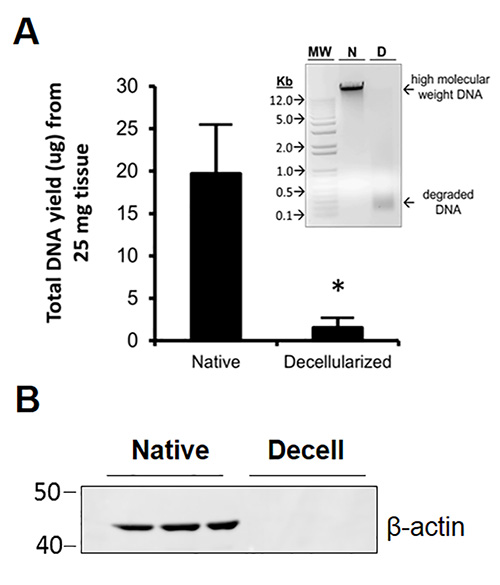
Figure 4. DNA and cellular protein is efficiently removed during decellularization. A) DNase treatment removes >90% (p<0.006) of DNA from the detergent-decellularized tissue. The small amount of remnant DNA that could be collected from the decellularized tissue was concentrated and run on a 0.8% agarose gel; the result (inset) showed that only low molecular-weight degraded fragments of DNA remained in the decellularized tissue. MW=molecular weight, N=native, D=decellularized. B) Western blot analysis showed that the representative cytoplasmic protein β-actin was not detectable in decellularized lung relative to native lung where this protein was abundantly detected. Reprinted with permission from reference19.
Click here to view larger image.

Figure 5. Fully assembled lung bioreactor. The bioreactor is assembled in a laminar flow hood to reduce the risk of contamination. A cell culture incubator water pan holds all bioreactor components and functions to collect spills if a leak occurs. The entire unit can be housed in an incubator. Levels of media in the various chambers should be determined empirically according to the researcher's needs.
Click here to view larger image.
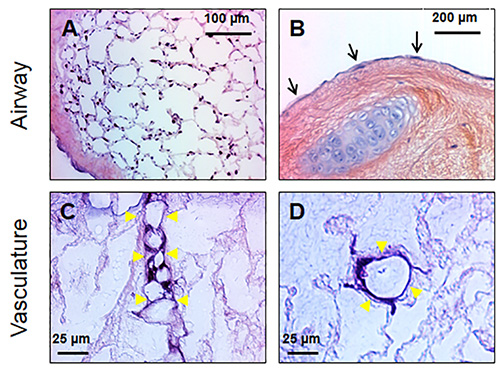
Figure 6. Bone marrow-derived mesenchymal stem cells (BMSC) and endothelial cells can be cultured within decellularized lung scaffolds. A) BMSC attach to the matrix scaffold of decellularized lung parenchyma and can be maintained with mechanical ventilation of culture media for extended periods; the panel shows a lung two weeks after seeding with BMSC. The cells line the alveolar septae creating the appearance of native lung. B) The lumenal surfaces of large airways serve as cell-attachment points as well. BMSC cultured for two weeks in the bioreactor grow along these surfaces and resemble simple squamous epithelium (black arrows). C) Seeding the vascular compartment of decellularized lungs with microvascular endothelial cells results in the recellularization of small vessels. Endothelial cells can be found attaching to the vascular basement membrane (yellow arrowheads). In some instances, the cell attachment points span the lumen (C). D) A cross-sectional view of a small vessel shows endothelial cells attached to the vessel basement membrane while maintaining an open vascular lumen.
Click here to view larger image.
