Rescue of NDV is a well-established procedure, routinely performed in the laboratories that have access to the complete cDNA of the virus. However, the intrinsic stochastic nature of the method makes it difficult to achieve 100% rescue efficiency. Monitoring the early steps of the process, specially the transfection efficiency and the infection with MVA-T7, helps identifying possible problems. Figure 5A shows standard transfection and transfection/infection efficiencies that are enough for a successful NDV rescue. After the rounds of amplification in avian-mammalian co-cultures and embryonated eggs, presence of rescued virus is detected by positive wells in the HA assay. NDV induces hemagglutination by linking erythrocytes and thus prevents their sedimentation in the bottom of the V-shaped well. Therefore, lack of HA activity can easily be distinguished by the formation of a red, round pellet of RBC (Figure 5B). Negative results in the HA assay can be due to an insufficient titer of virus in the allantoic fluid: a minimum titer of approximately 106 pfu/ml is required to have a positive HA result. Lower viral titers can still be detected by immunofluorescence assay (IFA) using specific anti NDV antibodies. A new passage on eggs for further amplification of the virus present in the first inoculated eggs is recommended in these cases.
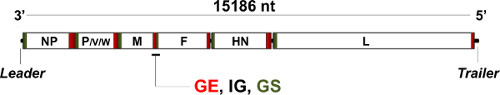
Figure 1. Genome organization of Newcastle disease virus (NDV). The non-segmented, single-stranded negative-sense RNA genome of NDV is 15,186 nucleotides (nt) long and encodes eight different polypeptides in six transcriptional units: NP, P, M, F, HN and L plus V and W proteins, originated after shifts in the reading frame of the P ORF during transcription. Noncoding, regulatory sequences flank the genome in its 3' (leader) and 5' (trailer) ends. Further noncoding gene end (GE), intergenic (IG) and gene start (GS) sequences regulate the viral polymerase activity in between the different ORFs. Expression levels of the different genes on the viral genome depend on their relative location respect to the 3' end (i.e. NP transcripts are the most abundant, L transcripts the least), effectively creating a gradient critical for the virus' life cycle1.
Figure 2. Basis of NDV rescue from cloned DNA. Schematic comparison between a natural (A) and the artificially-induced (B) cycle during the rescue process. (A) After attaching to the target cell, the virus releases its genomic content into the cytoplasm encapsidated in RNPs. The RNA polymerase transcribes the RNA- genome into the different mRNAs of the viral proteins and into a complementary, full length antigenomic RNA+ copy, which is template for the production of multiple copies of the original genome that will be incorporated into nascent virions. (B) During the rescue process, the components of the RNPs (NP, P and L proteins, as well as a full-length, usually modified antigenomic RNA) are provided by cDNA transfection. Upon reconstitution of the antigenomic RNPs in the cytoplasm of the transfected cell, they can produce the complementary, recombinant genomic RNPs and a whole cycle can then start anew, ending with the release of recombinant viruses.
Figure 3. T7-driven expression plasmids for the generation of recombinant NDV. (A) Full length viral cDNA: The full length pNDV-B1 ampicillin resistance plasmid driving expression of the lentogenic NDV strain Hitchner B1 antigenome under the T7 promoter (PT7) and the T7 terminator sequence (TT7) containing the Hepatitis Delta Ribozyme (HDR) cleavage. The pNDV-B1 was originally designed to carry foreign inserts between P and M genes5. Insertion of a new gene (XbaI site) requires the addition of the regulatory gene end (GE), intergenic (IG) and gene start (GS) sequences to allow its functional recognition by the viral polymerase. Expression efficiency of the transgene can be improved with the addition of a Kozak (K) sequence upstream the start codon32. The whole cassette introduced into the NDV cDNA needs to have a total number of nucleotides divisible by six to follow the "rule of six"33 that drives efficient replication of most paramyxovius genomes. (B) Protein expression plasmids: Viral NP, P, and L ORFs, flanked by the T7 promoter (PT7), the UTR of the encephalomyocarditis virus (EMC) and the T7 terminator (TT7) sequences were cloned into the pTM1 backbone ampicillin resistance vector5,36.
Figure 4. Plasmid-based reverse genetics techniques for the generation of recombinant Newcastle Disease Virus. (A) Viral rescue: A549 or HEp-2 cells are infected with the modified vaccinia virus Ankara expressing the bacteriophage T7 polymerase (MVA-T7). After viral infection, cells are co-transfected with the expression plasmid required for replication and transcription of the NDV viral genome (NP, P, and L), together with the full length NDV cDNA, under the T7 promoter. Twenty-four hours post-infection/transfection, mammalian cells are co-cultured with chicken or duck embryo fibroblasts (CEF and DEF, respectively). Three-to-four days after co-culture, 8-10 day-old chicken embryonated eggs are inoculated with the tissue culture supernatants of the co-culture cells for further amplification. Two-to-three days after incubation of the 8-10 day-old eggs at 37 °C, allantoic fluid from the eggs is harvested and analyzed for successful rescue of the recombinant virus by HA assay. Positive (+) rescue viruses are further characterized at genomic (RT-PCR) and protein (e.g. immunofluorescence assay (IFA) and western blot (WB) levels. Negative (-) HA results may be due to the lack of enough viruses to be detected by HA. Reinfection of fresh chicken embryonated eggs to amplify the virus is indicated. (B) Infection of chicken embryonated eggs: 8-10 chicken embryonated eggs are candled to mark the interface between the air sac and the allantoic cavity. With an insulin (1 ml) syringe, eggs are infected with the tissue culture supernatant inside the allantoic cavity, as indicated.
Figure 5. Representative results. (A) Transfection and MVA-T7 infection efficiencies: representative fluorescent (left) and bright (right) fields of A549 cells transfected with pCAGGS-GFP (top) or infected with MVA-T7 and transfected with pCITE-GFP (bottom) are illustrated. Constitutive GFP expression driven by pCAGGS is an indicator of transfection efficiency, whereas T7 promoter driven GFP expression by pCITE is a control for MVA-T7 infection and T7 polymerase activity. Cells were imaged at 24 hr post-transfection/post-infection (B) Hemagglutination assay (HA): Presence of viral particles in allantoic fluid induces hemagglutination, which is shown by a lack of pellet in the bottom of a V shaped well. Absence of virus allows the formation of a red pellet in the bottom of the well. Figure shows both the controls needed for any HA assay (negative, clean allantoic fluid; positive, an already characterized NDV sample), and the results of three unknown samples obtained after a rescue, with two positive (+) and one negative (-) result.
