The typical dependent measure in this test is the number of trials per stage to meet criterion (or 8 correct consecutive choices). Figure 4A shows the average number of trials to meet criterion at each stage in untreated C57BL/6J using the AST. As mice form an attentional set on the relevant cue dimension (odor or media), performance will improve, as indicated by a reduction in trials to meet criterion in successive intradimensional shifts, and the reversal stages and the extradimensional shift will require additional trials to meet criterion. It is important to counterbalance the number of animals that start on each of the two relevant dimensions (i.e., media or odor) as we have observed some difference at the simple discrimination stage reflected in an increased number of trials to criterion if odor is the relevant cue (Figure 4B). If the trials to reach criterion at the SD stage are significantly different between the two relevant cue dimensions (i.e., better performance when media is relevant as in Figure 4B), performance data for each mouse during subsequent stages can be normalized to its performance during the simple discrimination stage. As illustrated in Figure 4C, this normalization step reduces variability associated with learning the initially relevant cue and enhances visualization of improved performance on the subsequent IDS stages. Although group sizes of ~20 mice will be sufficient to allow analysis of raw data without normalization. Importantly, a significant increase in the trials to meet criterion during the extradimentional shift relative to the previous intradimentional shifts indicates is an important control measure of the test validity (Figure 4D & 4E). Prior to the ED shift stage, mice learn to pay attention to a single relevant cue dimension (odor or media) in order to locate the reward. The switch to a new relevant cue dimension, that was previously irrelevant, should require more trials to reach criterion, resulting in an ID/ED shift. Interestingly, analysis of the ID/ED shift within the raw data revealed a significant increase in the trials to reach criterion between IDS2 and ED (Figure 4D), and trend toward an increase between IDS3 and ED (data not shown, p<0.1) and an increase between the mean trials to reach criterion of IDS2 and IDS3 compared to ED (data not shown). Interestingly, a significant ID/ED shift was apparent between both IDS2 and IDS3 compared to ED when the data were normalized to performance at the SD stage (Figure 4E).
It is expected that approximately 70-80% of control mice will complete the entire task (20-30% drop out rate). Alternative dependent measures include average latency to make a choice as measured at each stage of the task (although we have found this to be highly variable between mice), total number of errors and number of “set-loss” errors at the reversal and extra-dimensional shift stages as defined by Lapiz-Bluhm et al.14 as the number of times an incorrect choice was made following three or more consecutively correct choices. An increased number of “set-loss” errors may indicate a perseverative behavior. Additionally, mice typically complete each stage of testing in less than 30 min (Figure 5), and the duration of time spent in each stage is a function of trials to reach criterion.
In a previous study using a similar paradigm as detailed here, neurotoxic lesions to the OFC or the mPFC in mice were shown to impair performance at the reversal and extra-dimensional shift stages respectively, validating the use of this test to determine functional deficits in the same regions as rat and non-human primate studies11. While the sequence of the tasks in the aforementioned study varies slightly to that shown here, it is expected that lesions or deficits in the OFC and mPFC will also result in impaired performance at R1 and EDS, as reflected by an increase in the number of trials to criterion at those stages. .
| Stage | Relevant | Examples | Relevant | Examples | ||
| Simple Discrimination (SD) | Medium | Felt (+) | Paper (-) | Odor | Nutmeg (+) | Rosemary (-) |
| Compound Discrimination (CD) | Medium | Felt (+)/Nutmeg | Paper (-)/Rosemary | Odor | Nutmeg (+)/Felt | Rosemary (-)/Paper |
| Felt (+)/Rosemary | Paper (-)/Nutmeg | Nutmeg (+)/Paper | Rosemary (-)/Felt | |||
| Reversal 1 (R1) | Medium | Paper (+)/Rosemary | Felt (-)/Nutmeg | Odor | Rosemary (+)/Paper | Nutmeg(-)/Felt |
| Paper (+)/Nutmeg | Felt (-)/Rosemary | Rosemary(+)/Felt | Nutmeg (-)/Paper | |||
| Intradimensional Shift (IDS) | Medium | Pompoms | Sequins (-)/Clove | Odor | Cinnamon (+)/Pompoms | Clove (-)/Sequins |
| (+)/Cinnamon | ||||||
| Pompoms (+)/Clove | Sequins (-)/Cinnamon | Cinnamon (+)/Sequins | Clove (-)/Pompoms | |||
| IDS2 | Medium | Pipecleaners | Googly eyes (-)/Ginger | Odor | Red thyme (+)/Pipecleaners | Ginger (-)/Googly eyes |
| (+)/Red thyme | ||||||
| Pipecleaners (+)/Ginger | Googly eyes (-)/Red thyme | Red thyme (+)/Googly eyes | Ginger (-)/ Pipecleaners | |||
| IDS3 | Medium | Ribbon (+)/ Vanilla | Metallic strips (-) /Lemon | Odor | Vanilla (+)/Ribbon | Lemon (-)/Metallic strips |
| Ribbon (+)/Lemon | Metallic strips (-) /Vanilla | Vanilla (+)/Metallic strips | Lemon (-)/Ribbon | |||
| R2 | Medium | Metallic strips (+)/Vanilla | Ribbon (-)/Lemon | Odor | Lemon (+)/Ribbon | Vanilla (-)/Metallic Strips |
| Metallic strips (+)/Lemon | Ribbon (-)/Vanilla | Lemon (+)/Metallic strips | Vanilla (-)/Ribbon | |||
| Extradimensional Shift (EDS) | Odor | Citronella (+)/Raffia | Anise (-)/Foam | Medium | Raffia (+)/Citronella | Foam (-)/Anise |
| Citronella (+)/Foam | Anise (-)/Raffia | Raffia (+)/Anise | Foam (-)/Citronella | |||
Table 1. Example combinations of odor and digging medium. The scents and digging media in these combinations have been validated in this study. Representative results obtained using these specific combinations are shown in Figure 2.
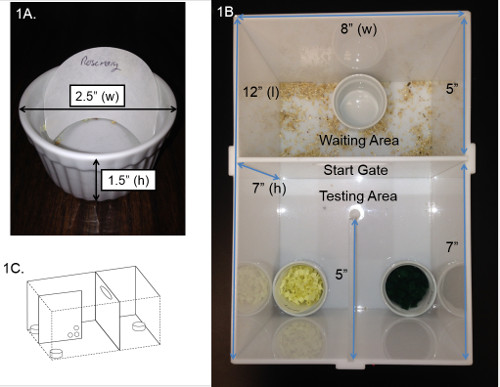
Figure 1. AST testing chamber. (A) Picture and dimensions of ramekin and example of filter paper placement. (B) Picture of the AST testing chamber adapted for mice and (C) a three dimensional diagram of the testing chamber illustrating placement of holes within the partition.
Figure 2. Digging medium. Examples of digging medium pairs used in this protocol as detailed in the materials list (ii.-xi.). Pots are filled approximately to the inner rim with digging medium.

Figure 3. Timeline for testing.
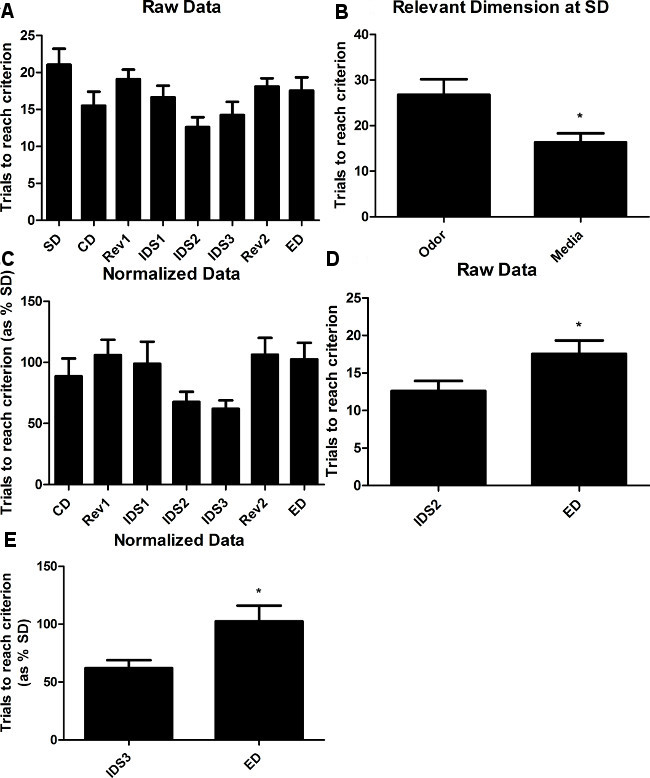
Figure 4. C57Bl/6J performance in the AST. (A) Average number of trials to meet criterion in the AST for male C57BL/6J mice. Data represent mean ± SEM (n=22 mice/group), p=0.013 for main effect of stage. (B) Average number of trials to reach criterion at the SD stage for distinct initial relevant cue dimensions. Data represent mean ± SEM (n=22 mice/group), *p=0.01. (C) Normalized data represented as a percentage of each mouse’s individual performance at the SD stage. Data represent mean ± SEM (n=22 mice/group), p=0.06 for main effect of stage. (D) ID/ED shift in trials to reach criterion between IDS2 and ED. Data represent mean ± SEM (n=22 mice/group), *p=0.03. (E) ID/ED shift in normalized data between IDS3 and ED. Data represent mean ± SEM (n=22 mice/group), *p=0.012.
