Methods for Cell-attached Capacitance Measurements in Mouse Adrenal Chromaffin Cell
Summary
After exocytosis, fused plasma membrane is retrieved through a process known as endocytosis. This mechanism reforms new synaptic vesicles for the next round of release. Individual endocytic events are captured and analyzed through the use of the cell-attached capacitance recordings in mouse adrenal chromaffin cells.
Abstract
Neuronal transmission is an integral part of cellular communication within the brain. Depolarization of the presynaptic membrane leads to vesicle fusion known as exocytosis that mediates synaptic transmission. Subsequent retrieval of synaptic vesicles is necessary to generate new neurotransmitter-filled vesicles in a process identified as endocytosis. During exocytosis, fusing vesicle membranes will result in an increase in surface area and subsequent endocytosis results in a decrease in the surface area. Here, our lab demonstrates a basic introduction to cell-attached capacitance recordings of single endocytic events in the mouse adrenal chromaffin cell. This type of electrical recording is useful for high-resolution recordings of exocytosis and endocytosis at the single vesicle level. While this technique can detect both vesicle exocytosis and endocytosis, the focus of our lab is vesicle endocytosis. Moreover, this technique allows us to analyze the kinetics of single endocytic events. Here the methods for mouse adrenal gland tissue dissection, chromaffin cell culture, basic cell-attached techniques, and subsequent examples of individual traces measuring singular endocytic event are described.
Introduction
Synaptic transmission is mediated by exocytosis of neurotransmitter-containing synaptic vesicles, and these vesicles must undergo local endocytic recycling within the nerve terminal to maintain neuronal communication in the long term. Given the essential role of synaptic transmission in the brain, understanding the molecular machinery that constitutes the synaptic vesicle cycle is an essential foundation towards a better comprehension in cellular communication as a whole. Among cell model systems, the adrenal chromaffin cell has provided some of the most definitive insight into the molecular machinery underlying synaptic vesicle recycling. Exocytosis, the final step in neurotransmitter release, has been immensely studied and examined through the use of the adrenal chromaffin cell1,2. In fact, most of the molecular players that orchestrate the formation, targeting, docking, and fusion of secretory granules have been identified due to application of diverse techniques in chromaffin cells1. Furthermore, by providing an opportunity to allow for single-vesicle resolution of the protein machinery involved in exocytosis, the chromaffin cell remains a powerful model to address the questions of vesicle fusion3.
Cell-attached capacitance measurements were first utilized in resolving single-vesicle fusion during exocytosis3. Exocytosis of vesicles as small as ~60 nm in diameter have been demonstrated to be detected by cell membrane admittance measurements with the patch clamp technique in the cell attached configuration4-7. Admittance is defined as a measure of how easily a circuit or device will allow a current to flow; it is the inverse of impedance. Thus, admittance measurements provide an understanding of the membrane capacitance. This is accomplished by the incorporation of the vesicular membrane into the plasma membrane; this incorporation reveals changes in surface area8. Each fusing vesicle causes a stepwise increase in membrane capacitance9,10. Additionally, this admittance measurement provides the membrane conductance and the fusion pore conductance during an exocytotic event3. As this technique has provided a unique tool at identifying single-vesicle kinetics during exocytosis, our lab has recently applied this concept to detect endocytosis of single vesicles11,12.
Our specific interest is clathrin-mediated endocytosis (CME), which has been considered as a fundamental housekeeping component in many cells13 and as a main pathway for synaptic vesicle endocytosis in neuronal terminals14,15. CME is known to be biologically important, however, its kinetics remain not well understood due to technical limitations in monitoring singular endocytic events. Given the similarities in exocytic mechanisms between chromaffin cells and neurons1, it is plausible that the fission mechanisms in chromaffin cells may likely apply to synaptic vesicle endocytosis in neurons. The cell-attached capacitance measurements have been utilized to monitor individual endocytic events and analyze the fission kinetics, which most methods are unable to resolve. In our cell-attached recordings, a sine wave at 20 kHz is superimposed over the holding potential, and the output current is separated into membrane conductance in one channel and membrane capacitance in the other channel from a two-phase lock-in amplifier16-18. From the changes in the membrane conductance and capacitance, one can calculate the kinetics of the fission-pore, which likely corresponds to the tubular membrane neck that connects the internalizing vesicle to the plasma membrane prior to vesicle pinch-off. Collectively, this technique gives us the opportunity to examine the regulatory mechanisms of vesicle fission during CME.
Protocol
NOTE: The entire procedure was conducted in accordance with the guidelines of the National Institutes of Health, as approved by the Animal Care and Use Committee of the University of Illinois at Chicago.
1. Solutions and Culture Media Preparations
- Keep all the solutions at -20 °C for up to six months. Keep the culture media at 4 °C for up to 3 months.
- Prepare 100 ml of culture media by mixing 1 ml of pen-strep solution and 1 ml of ITSX (Insulin-Transferrin-Selenium-Supplementation) to 100 ml with DMEM.
- Prepare enzyme solution by mixing 250 ml of DMEM, 2.5 ml of 100 mM CaCl2, and 2.5 ml of 50 mM EDTA.
- Prepare inactivation solution by mixing 225 ml of DMEM, 25 ml of FBS, 625 mg of albumin, and 625 mg trypsin inhibitor.
- Prepare a Locke’s Solution of 154 mM NaCl, 5.6 mM KCl, 5.0 mM HEPES, 3.6 mM NaHCO3, 5.6 mM glucose. Adjust the pH to 7.3 with NaOH.
- Prepare a poly-d-lysine (PDL) solution of 50 mg/ml. Use a final concentration of 1 mg/ml to coat coverslips.
- Prepare an extracellular solution of 140 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, 10 mM HEPES-NaOH, and 10 mM glucose. Adjust the pH to 7.3 with NaOH with an osmolarity of ~310 nmol/kg.
- Prepare a pipette solution of 50 mM NaCl, 100 mM TEACl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, and 10 mM HEPES. Adjust pH to 7.3 with NaOH with an osmolarity of ~290 nmol/kg.
2. Adrenal Gland Isolation
- Autoclave a pair of large and small dissection scissors and two pairs of forceps. Wear gloves during the whole process and place the dissecting tray on an ice bucket.
- Use mice taken at Day 0-3. Euthanize animals with a pressurized CO2 tank followed by decapitation.
- Utilize the smaller set of scissors to cut down the back of the pup following the spine from cervical to caudal region. Be sure to cut superficially under the skin and not pierce into the body cavity.
- Next, peel skin away from spine so that musculature is exposed to have a clear view of the spinal column. From here, make a transectional cut at the cervical spinal cord and then make two parallel cuts along the sides of the spine.
- With one pair of forceps holding the main body of the animal, use the other pair of forceps to grab spinal column and peel back spine till kidneys are exposed.
- At this moment, visualize the adrenal glands on the top of the kidneys. Then carefully place the two glands from each animal into a conical tube filled with Locke’s solution.
NOTE: Sometimes the glands will stick to the forceps. If this is the case, gently use another pair of forceps to aid in the removal of the glands from the forceps and into the Locke’s solution. The glands can be kept in this condition for up to 2 hr.
3. Tissue Digestion
- In the meantime, bubble the enzyme solution with 5% CO2 + 95% O2 for 15 min prior to use. Ensure that the equilibrated solution turns from a bright pink color into pinkish/orange color.
- Add the papain to the freshly bubbled enzyme solution at a final concentration of 20-25 unit/ml. Incubate each pair of glands from each animal for 40-60 min at 37 °C in the enzyme solution with papain.
- Add 75% of inactivation solution to each tube and incubate at 37 °C for another 10 min. This incubation inactivates the enzyme activity of papain.
- After the 10 min incubation with the inactivation solution, gently transfer the glands into a newly labeled tube and then wash the glands 3x with culture media.
4. Cell Dissociation and Plating
- After washing, place the two glands into 200 µl of the culture media and then gently titrate the glands up and down through the pipette tip with a 200 µl pipette until there is no longer a large piece of tissue in the solution.
NOTE: When titrating, maintain glands at the bottom of the tube with a downward force from the 200 µl pipette tip and engage the tissue gently and slowly. Be certain to use slow, continuous strokes, never pipetting fast or abrupt. - Add an additional 250 µl culture media to bring the total volume of culture media up to 450 µl.
- Plate 60 µl of this cell-containing solution onto each of seven 12 mm PDL-pre-coated coverslips, which are placed in a 60 x 15 mm culture dish.
- Once plated, place the dish into the incubator that is set to 37 °C and maintained with a steady flow/supply of 5% CO2 for 30 min to ensure an environment for cell viability. After incubation, gently add 5 ml of pre-warmed culture media into the culture dish and return the dish back into the incubator for 24 hr prior to cell-attached recordings.
NOTE: Typically, the cells can be used for electrophysiological recordings for up to 4 days.
5. Cell Identification and Gigaohm Seal Formation
- Place the 12 mm coverslip into extracellular solution. Since the whole adrenal glands are used for cell preparation, mix the chromaffin cells (typically very bright with a brownish/beige coloring and a near perfect circular form (Figure 1) in culture with cortical cells (smaller and dimmer than chromaffin cells).
- Pull the patch pipettes in four stages using a programmable P-97 puller. Dip the pipette tip into a melted wax, which will help to reduce the capacitance of the glass and then fire polish the tip to “blow-away” this wax from inside of the pipette tip.
NOTE: When filled with patch pipette solution, patch pipettes typically have a resistance of ~2 MΩ. - Approach the patch pipette close to the cell within several microns. To achieve a cell-attached configuration, look for any slight cell deformation during the approach of the patch pipette closer to the cell. Apply gentle suction until a GΩ seal is achieved.
6. Cell-Attached Capacitance Recordings
- Once a GΩ seal is revealed, make cell-attached recordings through the use of an EPC-7 plus patch-clamp amplifier and a two-phase analog lock-in amplifier (see attached table of equipment). Apply a sinewave with amplitude of 50 mV (r.m.s.) and frequency of 20 kHz from the lock-in amplifier.
- Set the output filter of the lock-in amplifier at 1 msec time constant and 24 db. Set the phase of the lock-in amplifier such that transient capacitance changes produced by gentle suction pulses appear only in the patch capacitance (Im) trace with no projection into the patch conductance (Re) trace (Figure 2).
NOTE: Please see the details for the calibration of the system in reference 19. - Identify typical capacitance changes by “downward” steps (Figure 3).
NOTE: A solid recording will be viewed as a “staircase” type resemblance with many downward steps, which indicates internalizing single vesicles. The patching process serves as a mechanical stimulation since action current can be typically recorded from most cells. - Name files as individual folders including date: mm/dd/year and each cell-attached recording as its own individual numbering (xx) within that file.
Representative Results
The cell viability and the quality of gigaohm seal are critical in determining the quality of the cell-attached capacitance recordings. Therefore, it is critical to procure an effective and efficient cell culture prior to electrophysiological recordings, and typical viable cells are illustrated in Figure 1. Practice and time will be helpful in achieving a gigaohm seal with high quality. If one can clearly see cell deformation when the patch pipette is approaching the cell as described in protocol Step 5.3, there is a better chance in obtaining a high-quality seal. Figure 3 demonstrates a typical recording of membrane capacitance with multiple downwards steps associated with single vesicle endocytosis.
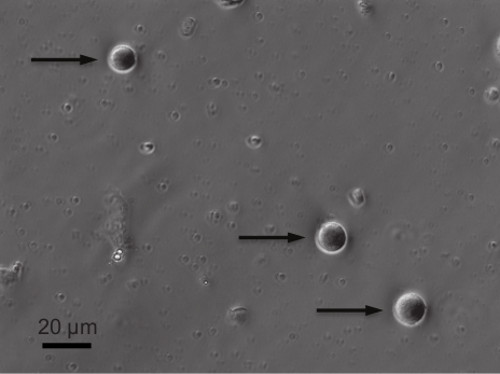
Figure 1. Examples of viable mouse adrenal chromaffin cells 24 hr after cell culture. Arrows point to 3 viable chromaffin cells. Please click here to view a larger version of this figure.
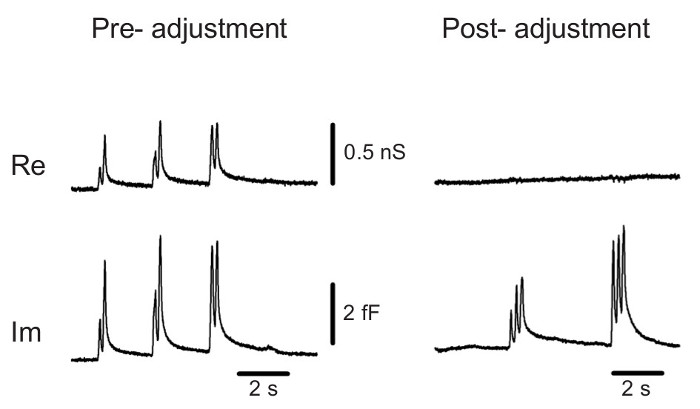
Figure 2. Phase adjustment in the cell-attached recordings. With the initial phase setting, gentle suctions cause transient changes in both capacitance (Im) and conductance (Re) traces, and the projections in Re trace are cancelled out by a 23° phase shift. Please click here to view a larger version of this figure.
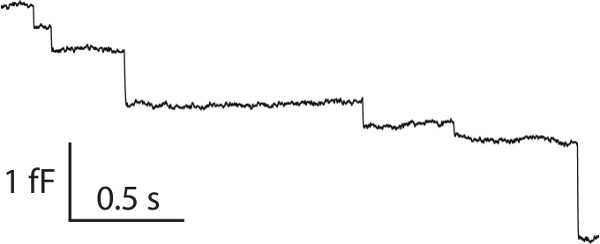
Figure 3. A typical cell-attached capacitance recording with multiple downward steps, associated with single vesicle endocytosis. Please click here to view a larger version of this figure.
Discussion
Cell-attached capacitance measurements require several critical steps in order to successfully obtain recordings with high quality: 1) viable and healthy cells prepared from adrenal glands; 2) PDL coating of the coverslips; 3) gigaohm seal formation; 4) noise level of the system; and 5) phase correction.
For animal surgery, modifications one can potentially make is to adjust the surgical approach to best suit dexterity and to prevent damage during dissection. Additionally, sufficient practice on locating and removing the adrenal glands is imperative as cells that will be utilized for electrophysiological recordings come exclusively from the medulla of the adrenal gland. For enzyme digestion, it is critical to equilibrate the enzyme solution by bubbling the solution with 5% CO2 + 95% O2. Be certain that the tubing connected to the cylinder is sufficient and tight with no leaks. Additionally, be sure that the end in solution is properly placed during the entire 15 min bubbling time; this can be achieved with placing a 20 G needle end that has been blunted at the end of the tubing so as to direct air flow appropriately. Moreover, additional time for this step will not hurt so long as the airflow is not so powerful as to overflow the solution in the tube. Cell titration is another critical component that will take practice. The majority of the cells can be damaged if over-titrated; conversely, if titration is not enough, there will be very few single isolated cells dissociated from the tissues. To effectively titrate, utilize a 200 µl pipette tip to pipette up and down 7-8x, making sure the glands are gently passed through the 200 µl pipette tip as you titrate.
For PDL coating, one can always extend the incubation time of the PDL on the coverslips up to 3 hr to ensure sufficient coating of coverslips. The PDL coating is critical for chromaffin cells to attach to and grow on the coverslips after cell plating. If PDL coating is not sufficient, most of the cells will be detach from the coverslip, which will make the gigaohm seal formation difficult.
Cell-attached recordings can always be modified and improved, as the patching technique is a dedicated process. Gentle and subtle suction is critical in facilitating the gigaohm seal formation when the patch pipette and cell are in contact. Too much suction will destroy the patch tip-cell contact and in extreme situations, the cells can be sucked into the patch pipette. This will be very easily observed on the oscilloscope by changing into a whole cell configuration or by a very minimal resistance returning to a very large one; practice is of the utmost importance to obtain a high quality gigaohm seal formation.
While high seal resistance is critical to minimize the noise level in the cell-attached recordings, the following two steps are also important to reduce the noise level to resolve single endocytic events: (1) In the patch pipette solution, TEACl is utilized to block voltage-gated K+ channels; (2) the pipette tip is coated with sticky wax and the wax inside the pipette tip can be removed by heat polishing.
While the phase adjustment detailed in Protocol step 6.2 is critical to set up the initial phase for the recording, this step may not be ideal. Therefore, it is important to perform phase correction during fission-pore analysis for individual events. At a resting membrane potential of -65 mV, whole-cell recordings show that a typical mouse chromaffin cell has an input capacitance of 5 pF and an input resistance of 500 MΩ, which calculates the membrane conductance of mouse chromaffin cells as ~0.4 pS/fF. This implies that an endocytic event with a capacitance size of 1 fF will result in a net loss of membrane conductance of ~0.4 pS. This value is negligible compared to a typical 50-100 pS membrane conductance change associated with endocytic fission-pore closure in cell-attached recordings. Therefore, we are confident that our phase correction allows us to adjust the baseline of membrane conductance such that the pre and post fission-pore closure is at the same level; this process may bring <1% error into our data analysis.
In summary, the protocol described here demonstrates how cell-attached capacitance recordings can be utilized to monitor single vesicle endocytosis using mouse chromaffin cells as the model system. This technique allows one to analyze the regulatory mechanisms for vesicle fission during endocytosis.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work is supported by a National Science Foundation award (1145581) to LWG.
Materials
| Name of Material/ Equipment | Company | Catalog Number | Comments/Description |
| Poly-D-Lysine | Sigma | P0899 | |
| DMEM | 15066024 | Keep out of UV | |
| Dulbecco’s Modified Eagle Medium | Life Technologies | ||
| Cover Glass | Carolina Biological | 633029 | 12mm |
| Penicillin Streptomycin | Life Technologies | 15140122 | 100mL |
| Insulin-Trans-Sel-X | Life Technologies | 51500056 | Only thaw on ICE! |
| Papain | Worthington | 39S11614 | |
| EPC-7 plus patch amplifier | HEKA | ||
| BNC-2090 data acquisition board | National Instruments | ||
| Igor data acquisition software | Wavemetrics | ||
| P-97 pipette puller | Sutter Instruments | ||
| Microforge | Scientific Instruments | ||
| Borosilicate glass capillaries | Sutter Instruments | B150-110-10 | Outer diameter-1.5 mm |
| Inner diameter 1.10 mm | |||
| Length- 10 cm |
References
- Rettig, J., Neher, E. Emerging roles of presynaptic proteins in Ca++-triggered exocytosis. Science. 298, 781-785 (2002).
- Gong, L. W., et al. Phosphatidylinositol phosphate kinase type I gamma regulates dynamics of large dense-core vesicle fusion. Proceedings of the National Academy of Sciences of the United States of America. 102, 5204-5209 (2005).
- Lindau, M., Alvarez de Toledo, G. The fusion pore. Biochimica et Biophysica Acta. 1641, 167-173 (2003).
- Albillos, A., et al. The exocytotic event in chromaffin cells revealed by patch amperometry. Nature. 389, 509-512 (1997).
- Lollike, K., Borregaard, N., Lindau, M. Capacitance flickers and pseudoflickers of small granules, measured in the cell-attached configuration. Biophys J. 75, 53-59 (1998).
- Lollike, K., Borregaard, N., Lindau, M. The exocytotic fusion pore of small granules has a conductance similar to an ion channel. The Journal of cell biology. 129, 99-104 (1995).
- Dernick, G., Gong, L. W., Tabares, L., Alvarez de Toledo, G., Lindau, M. Patch amperometry: high-resolution measurements of single-vesicle fusion and release. Nat Methods. 2, 699-708 (2005).
- Neher, E., Marty, A. Discrete changes of cell membrane capacitance observed under conditions of enhanced secretion in bovine adrenal chromaffin cells. Proceedings of the National Academy of Sciences of the United States of America. 79, 6712-6716 (1982).
- Gong, L. W., de Toledo, G. A., Lindau, M. Exocytotic catecholamine release is not associated with cation flux through channels in the vesicle membrane but Na+ influx through the fusion pore. Nature Cell Biology. 9, 915-922 (2007).
- Gong, L. W., Hafez, I., Alvarez de Toledo, G., Lindau, M. Secretory vesicles membrane area is regulated in tandem with quantal size in chromaffin cells. The Journal of Neuroscience : the Official Journal of the Society for Neuroscience. 23, 7917-7921 (2003).
- Yao, L. H., et al. Actin polymerization does not provide direct mechanical forces for vesicle fission during clathrin-mediated endocytosis. The Journal of Neuroscience : the Official Journal of the Society for Neuroscience. 33, 15793-15798 (2013).
- Yao, L. H., et al. Synaptotagmin 1 is necessary for the Ca2+ dependence of clathrin-mediated endocytosis. The Journal of Neuroscience : the Official Journal of the Society for Neuroscience. 32, 3778-3785 (2012).
- McMahon, H. T., Boucrot, E. Molecular mechanism and physiological functions of clathrin-mediated endocytosis. Nature reviews. Molecular Cell Biology. 12, 517-533 (2011).
- Murthy, V. N., De Camilli, P. Cell biology of the presynaptic terminal. Annu Rev Neurosci. 26, 701-728 (2003).
- Granseth, B., Odermatt, B., Royle, S. J., Lagnado, L. Clathrin-mediated endocytosis is the dominant mechanism of vesicle retrieval at hippocampal synapses. Neuron. 51, 773-786 (2006).
- Rosenboom, H., Lindau, M. Exo-endocytosis and closing of the fission pore during endocytosis in single pituitary nerve terminals internally perfused with high calcium concentrations. Proceedings of the National Academy of Sciences of the United States of America. 91, 5267-5271 (1994).
- MacDonald, P. E., Eliasson, L., Rorsman, P. Calcium increases endocytotic vesicle size and accelerates membrane fission in insulin-secreting INS-1 cells. Journal of Cell Science. 118, 5911-5920 (2005).
- Zhao, Y., Fang, Q., Straub, S. G., Lindau, M., Sharp, G. W. Hormonal inhibition of endocytosis: novel roles for noradrenaline and G protein G(z). J Physiol. 588, 3499-3509 (2010).
- Debus, K., Lindau, M. Resolution of patch capacitance recordings and of fusion pore conductances in small vesicles. Biophys J. 78, 2983-2997 (2000).
