We have constructed a library of Hsp104 variants randomized in the middle domain and screened it for suppression of TDP-43 toxicity. The library was transformed and plated onto glucose and galactose plates (Figure 2) to assess the stringency of the screen. Single colonies were selected and the strains were counter selected using 5-FOA to eliminate the Hsp104 plasmid. These strains were then assessed to confirm that toxicity was due to TDP-43 alone, without the Hsp104 variants. Spotting assays of a subset of the variants selected in the initial screen showed that of the 4 colonies selected, 2 displayed Hsp104-mediated TDP-43 toxicity suppression, 1 was a false positive, and 1 displayed enhanced toxicity following 5-FOA treatment (Figure 3). The 2 true hits were then sequenced by colony PCR to identify the middle domain mutations (Figure 4). Once these mutations were identified, the Hsp104 mutation was constructed afresh in the parental Hsp104 plasmid by site-directed mutagenesis to confirm toxicity suppression. Variants selected using these methods were later confirmed to suppress aggregation in yeast, clear preformed aggregates in biochemical assays, and suppress dopaminergic neurodegeneration in a C. elegans model of PD17.
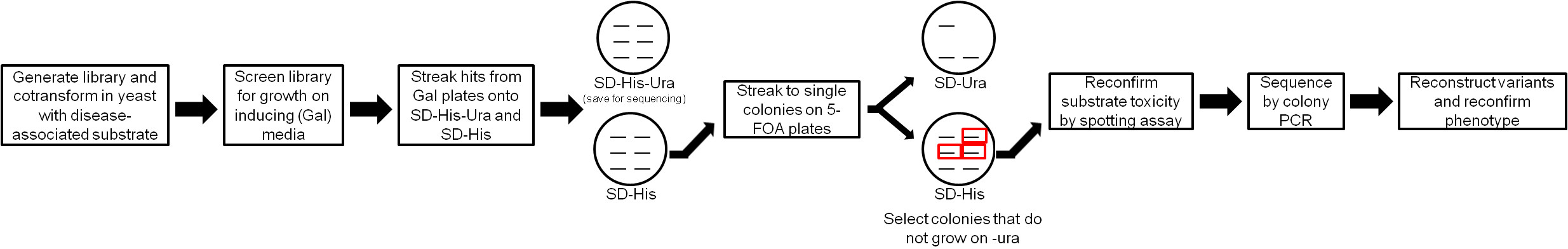
Figure 1: Flow-chart for isolating potentiated Hsp104 variants. Hsp104 libraries (URA3 marker, GAL1 promoter) are transformed in yeast containing the disease substrates (HIS3 marker, GAL1 promoter) and screened for toxicity suppressors by plating on inducing media. Potential hits are then screened again using a 5-FOA counterselection step to eliminate nonspecific toxicity suppressors. Variants selected in this second step are then sequenced by colony PCR. Please click here to view a larger version of this figure.

Figure 2: Screening for toxicity suppressors. Yeast cotransformed with pAG303GAL-TDP-43 and the pAG416GAL-Hsp104 library were plated onto glucose (repressing) or galactose (inducing) media. TDP-43 is highly toxic, so very few Hsp104 variants are capable of suppressing this toxicity, and this screen is very stringent. Single colonies are selected as hits from the galactose plate for the 5-FOA secondary screen. Both large and small colonies are typically obtained, but we have not observed any trends that dictate how colony size correlates with activity.
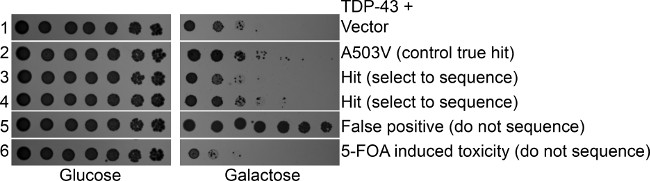
Figure 3: 5-FOA secondary screen. Yeast treated with 5-FOA to counter select for the Hsp104 plasmid were assessed by spotting assay. Strains were grown in SRaff-His media, serially diluted 5 fold, and spotted in duplicate onto SD-His and SGal-His plates. Hsp104A503V is a true hit that we have previously verified and show here as a control along with vector alone17. Rows 3 and 4 are library hits that retain TDP-43 toxicity following loss of the Hsp104 suppressor. Row 5 shows a false positive, where TDP-43 is no longer toxic, so a nonspecific toxicity suppressor is present. Row 6 shows a strain that is more toxic than TDP-43 typically is, possibly due to 5-FOA toxicity. This strain could still be sequenced but may not be a valid hit.
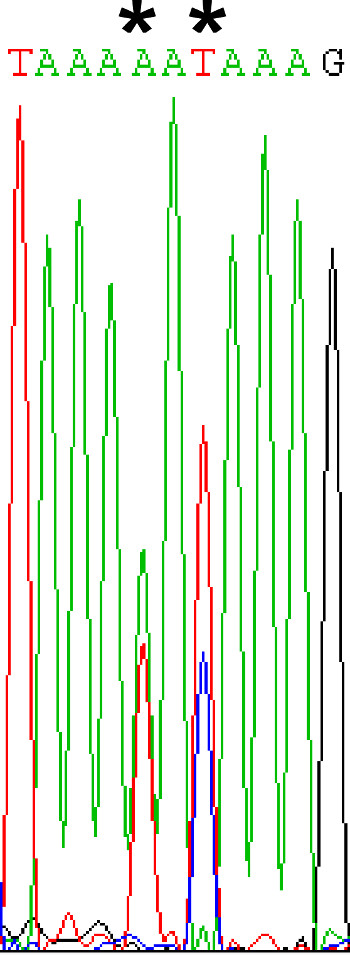
Figure 4. Analyzing sequencing results. Selected yeast often contains multiple plasmids. Therefore, following sequencing of the colony PCR products, care must be taken to ensure any mutations are not overlooked in the chromatograms. In this chromatogram, two potential mutations (denoted by *) could be overlooked. In the first instance, there is a mixture of A and T and in the second, a mixture of T and C.
