FRET flow cytometry enables sensitive, quantitative, and rapid detection of seeding activity from recombinant or biological samples. Assay setup is facile: monoclonal-derived stable cell lines expressing tau-RD-CFP/YFP are transduced with seed material, incubated for 24-48 hr, and subjected to flow cytometry analysis (Figure 1A). In the absence of seeds, biosensor cells maintain tau in a soluble, monomeric form (Figure 1B). In the presence of seeds, however, biosensor cells convert tau into an aggregated state (Figure 1C), producing a FRET response that is detected on a per-cell basis with flow cytometry. Quantitative assessment shows that biosensor cells respond to femtomolar concentrations of seed material and that seeding activity can be effectively measured across three orders of magnitude (Figure 1D).
Gating parameters were established to ensure detection of a true FRET signal, even at low seed concentrations. First, standard gating methods are used to isolate the cell population (Figure 2A) and single cells (Figure 2B). False FRET signal can arise from direct activation of YFP by the 405 nm laser. Thus, a False FRET gate is drawn from a YFP single-positive cell population to exclude contaminating signal (Figure 2C). Last, the FRET gate is defined from unseeded CFP/YFP dual-positive cells (Figure 2D). A gate is drawn near the slope of the population that extends upward and leftward away from it. Unstimulated cells should have a background FRET value set to approximately 1%, and FRET-positive cells will shift into this gate in a dose-dependent manner (compare Figure 2E and 2F). Multiple parameters are taken into account with FRET flow cytometry, including: percent positivity (i.e., the number of FRET-positive cells per total cell count) and median fluorescence intensity (i.e., the degree to which a FRET response is produced in a FRET-positive cell). Integrated FRET density is the preferred outcome measure, as it is the product of these two variables and wholly represents the degree of seeding activity induced by any given sample.
FRET flow cytometry is compatible with P301S tauopathy mouse-derived brain lysates, even at young ages, as displayed in Figure 3. Following microdissection of four different brain regions, FRET flow cytometry was used to measure seeding activity from brainstem (Figure 3A), neocortex (Figure 3B), frontal lobe (Figure 3C), and hippocampus (Figure 3D) in P301S mice over a range of ages. In each region, seeding activity increased with age and appeared at just 1.5 months. However, tau knockout mice (“KO”) >12 mos never displayed seeding activity. Compared to histological analyses performed within the same animals, this is six weeks sooner than the appearance of any other marker of tau deposition (Figure 3E), including markers of conformationally-aberrant tau (MC1), hyperphosphorylated tau (AT8 and PG5), and amyloid conformation (ThioflavinS). From a mechanistic view, the proximal detection of tau seeding activity suggests seeds as a causal mediator of tauopathy onset and/or progression. From an experimental view, this suggests that analysis of seeding activity may be an ideal supplement to standard outcome measures of tau deposition, given its early and robust appearance.
In addition to transgenic mice, FRET flow cytometry is compatible with human brain lysates and can readily detect tau aggregates isolated from Alzheimer's disease subjects (Figure 4). When frozen human brain tissue is homogenized in the same manner as described here for P301S brain lysates, seeding activity is robustly detected from all samples derived from AD subjects. In contrast, seeding activity is never detected from lysates of age-matched or Huntington's disease control subjects. This emphasizes the specificity of the assay for tau aggregates.
While the focus of this article is sensitive detection of tau seeding activity using liposome-mediated delivery of aggregates into HEK 293T biosensor cells, more physiological cell culture paradigms can also be performed with FRET flow cytometry to assess basic cellular uptake mechanisms. For example, primary neuronal cultures can be infected with lentivirus expressing the tau-RD-CFP and tau-RD-YFP constructs at time of plating. At DIV4, aggregation is absent in untreated cells (Figure 5A) but detectable in neurons treated with seed-containing material (Figure 5B). Importantly, phospholipids are excluded in these experiments, permitting the investigation of physiological and pathophysiological seeding mechanisms. Thus, application of tau fibrils leads to neuronal HSPG-mediated uptake and stimulates aggregation in a dose-dependent manner that can be quantified with FRET flow cytometry (Figure 5C). Additionally, the assay can be employed for therapeutic evaluation of tau uptake and seeding blockade in vitro using HEK 293T tau biosensor cells. Even in the absence of phospholipids, both recombinant fibrils (Figure 5D) and P301S brain lysates (Figure 5E) induce tau aggregation. Pre-incubation of these tau seeds with heparin (previously shown to be a potent inhibitor of HSPG-mediated tau uptake), however, eliminates their seeding capacity. This experimental setup could be applied to any drug (e.g., antibodies, small molecules, etc.) and would enable rapid evaluation of uptake and/or seed-modifying therapeutics.
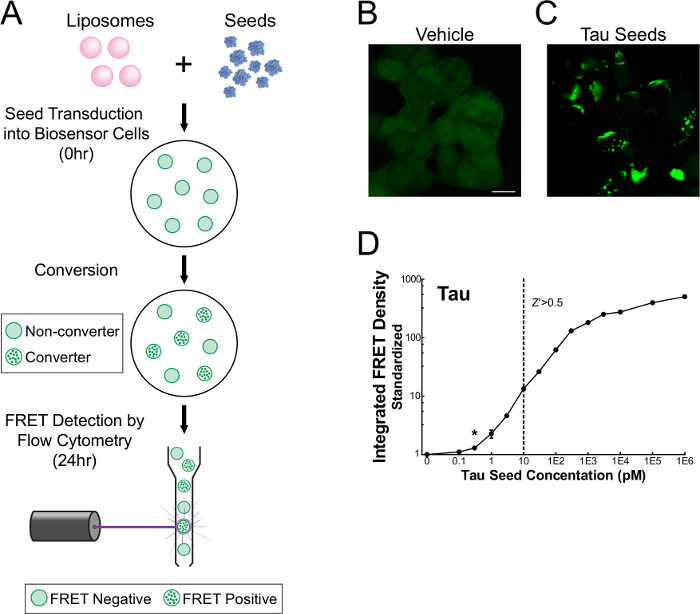
Figure 1. FRET flow cytometry sensitively detects tau seeding activity16. Monoclonal HEK 293T cells expressing tau-RD-CFP/YFP were transduced with recombinant or biological samples, incubated for 24-48 hr, and analyzed on a single cell basis using flow cytometry (A). Unstimulated cells maintain tau-RD in a monomeric state (B), whereas cells treated with seed-containing material display prominent inclusions (C). Quantitative assessment (mean ± S.E.M.) of seeding activity shows that FRET flow cytometry is sensitive to femtomolar concentrations (monomer equivalent) of recombinant seed material and detection spans three orders of magnitude (D). Monomer equivalent represents the total amount of protein contained within the fibrillization reaction and does not correct for the incomplete incorporation of monomer into aggregated material. Thus, the concentration of actual aggregates (seeds) must be less than or equal to its 'monomer equivalent'. *Modified from Holmes and Furman et al.16 Please click here to view a larger version of this figure.
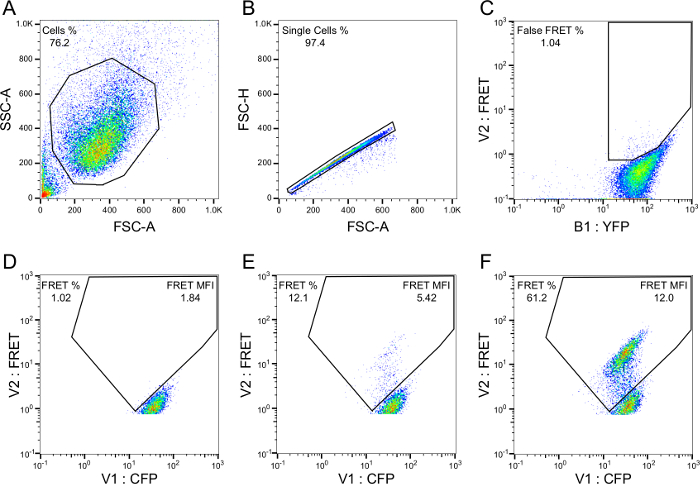
Figure 2. Gating strategy for FRET flow cytometry. Cell population (A) and singlet/doublet (B) gates are drawn with standard flow cytometry methodology. A false FRET gate (C) is drawn from YFP single-positive cells to eliminate YFP bleedthrough into the FRET filter. A FRET gate (D) is constructed from empty liposome-treated cells, such that background FRET is ≥1%. A population shift into the FRET gate appears following treatment with seed-positive material (E) and the shift becomes increasingly prominent with higher amounts of seed material (F). Final readouts include: percent FRET positivity, median fluorescence intensity (MFI) of FRET-positive events, and the integrated FRET density (Integrated FRET Density = Percent positive cells * MFI). Please click here to view a larger version of this figure.
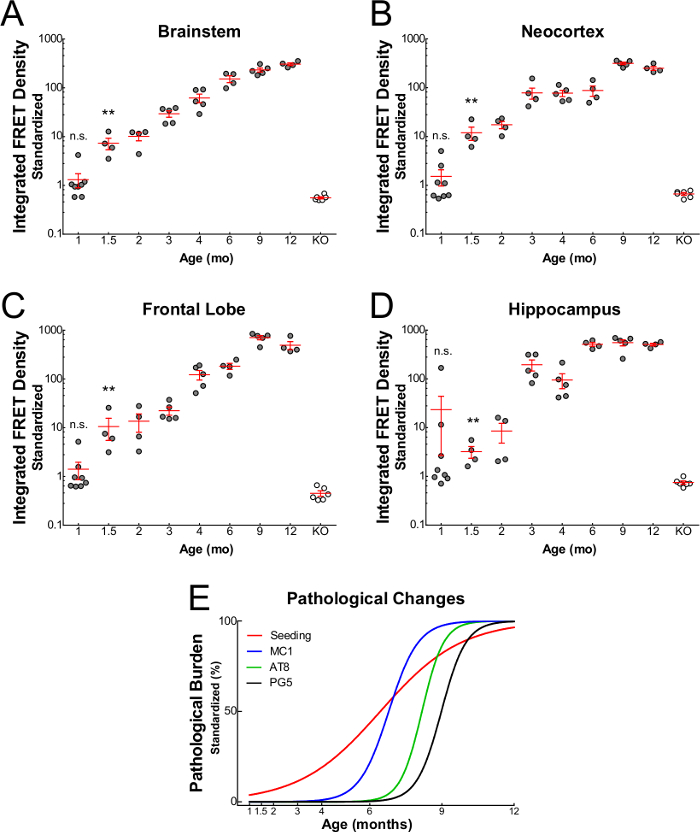
Figure 3. Progression of tau seeding activity in P301S mice16. Seeding activity (mean ± S.E.M.) from brain homogenates of P301S mice is apparent at 1.5 months in brainstem (A), neocortex (B), frontal lobe (C), and hippocampus (D). However, seeding activity is never observed in tau knockout mice (>12 months). Seeding activity increases with age and precedes the appearance of other common histological markers, including MC1, AT8, and PG5 (E). *Reprinted with permission from Holmes and Furman et al.16 Please click here to view a larger version of this figure.
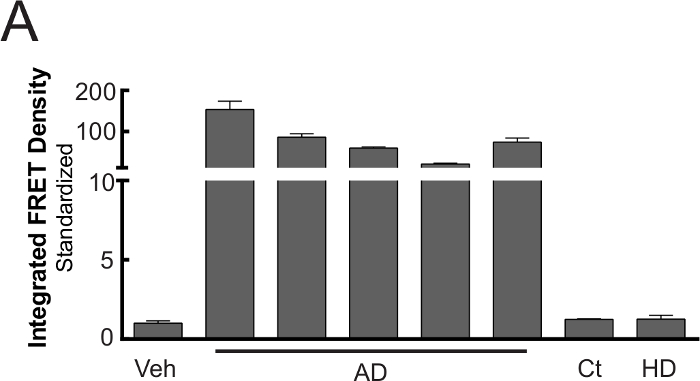
Figure 4. Compatibility of FRET flow cytometry with human biological samples16. When tau-RD-CFP/YFP biosensor cells are transduced with 20 µg human brain lysate, robust seeding (mean ± S.E.M.) occurs in all tested Alzheimer's disease brains, whereas age-matched and Huntington's disease control lysates lack seeding activity. *Modified from Holmes and Furman et al.16 Please click here to view a larger version of this figure.
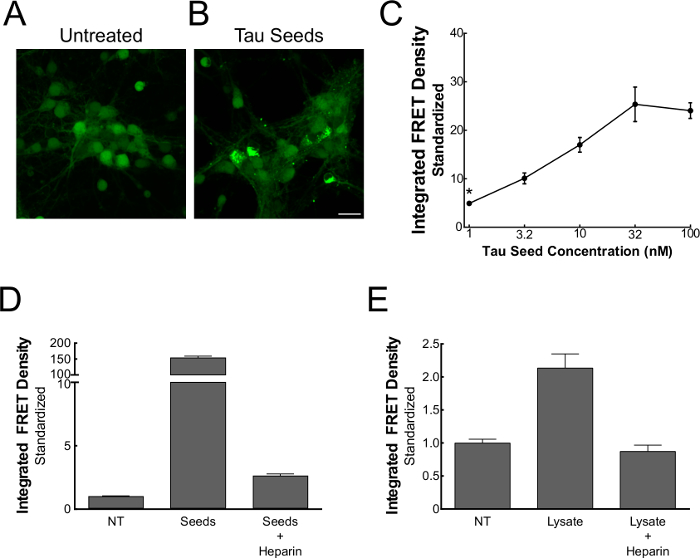
Figure 5. Alternative applications for FRET flow cytometry16. Primary neurons transduced with lentivirus encoding tau-CFP and tau-YFP constructs. In the absence of exogenous seeds there is no apparent aggregation (A), but there is in the presence of seeds (B). Neuronal biosensors respond dose-dependently to recombinant tau seeds, even in the absence of phospholipid-mediated delivery (C). Pre-incubation of recombinant (D) or biological (E) seed sources with heparin, an inhibitor of HSPG-mediated tau seed uptake, depletes FRET responsiveness of HEK 293T biosensor cells. Quantitative assessment displays (mean ± S.E.M.). *Modified from Holmes and Furman et al.16 Please click here to view a larger version of this figure.

Table 1: Cell lines used with FRET flow cytometry. HEK 293T cells are used for flow cytometry setup. CFP single-positive cells are used for compensation (*). YFP single-positive cells are used to eliminate false FRET signal due to direct activation of YFP by 405 nm excitation. CFP/YFP dual-positive cells are FRET-compatible and serve as the biosensor cells. Please click here to view a larger version of this table.
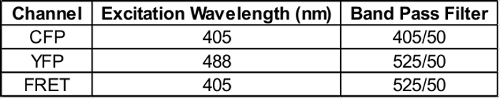
Table 2: Flow cytometer laser and filter settings. Please click here to view a larger version of this table.
