To prevent cell death induced by the hyperactive MAPK signaling, the kinase inactive versions of YDA (DNyda) and MPK6 (DNmpk6) were used in the co-expression assay in tobacco cells. Neither the interaction between DNyda and DNmpk6 revealed by BiFC nor CFP-BASL itself generated uneven distribution pattern (Figure 3A-B). However, when CFP-BASL was introduced into the DNyda-DNmpk6 interaction pair, both CFP and YFP signals were redistributed to a highly polarized manner (Figure 3C). This redistribution suggests that BASL interacts with YDA and MPK6 to spatially re-organize the MAPK signaling pathway in plant cells. The phosphorylation status of BASL has differential impacts on the polarity degree of the MAPK signaling: a phospho-mimicking version of BASL generated very strong polarity, whilst a phospho-deficient version showed weaker activity6. These data supported our working model of a positive feedback regulation between BASL and the YDA-MAPK pathway in generating cell polarity6.
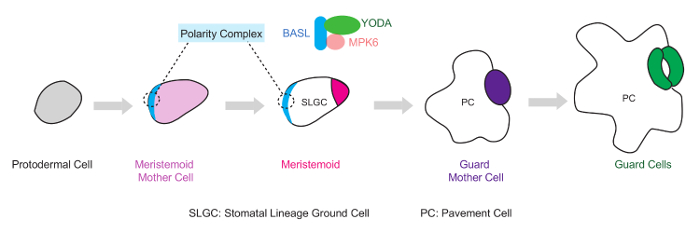
Figure 1. Diagram of the BASL-YDA-MPK6 Polarity Complex During Stomatal Asymmetric Cell Division (ACD) in Arabidopsis. In the leaf epidermis, the stomatal linage initiates from the protodermal cells, which expand to become ACD precursor cells, the Meristemoid Mother Cells (MMCs). A MMC undergoes an ACD to create one Meristemoid and one SLGC (stomatal lineage ground cell), which may divide and eventually differentiate into stomatal guard cells and pavement cells, respectively. At the cell cortex of the ACD precursor cells (MMCs), BASL recruits two components of a MAPK pathway, the MAPKKK YDA and the MAPK MPK6, to form a polarity complex and regulate stomatal ACD. Please click here to view a larger version of this figure.
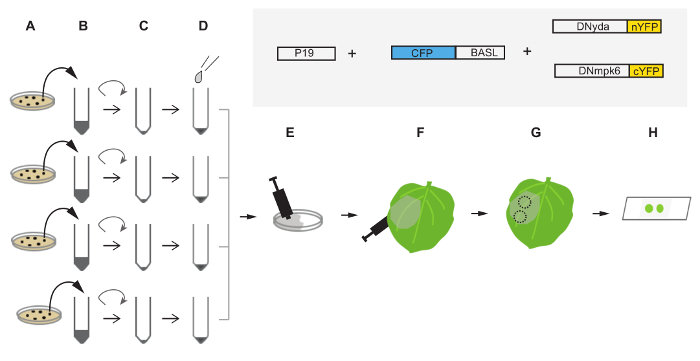
Figure 2. Diagram of the Injection Procedure in Tobacco Leaf Epidermis. (A) Transform the plasmids harboring CFP-BASL, DNyda-nYFP, DNmpk6-cYFP and P19 into Agrobacterium tumefaciens GV3101 and grow them on respective selection plates (from top to bottom in A). (B) Pick up colonies from (A) to inoculate liquid culture for overnight growth. (C) Harvest cells by spinning and decant the supernatant. After briefly pipetting and washing the pellets, spin down the cells again. (D) Re-suspend and dilute the cell pellets with the infiltration solution (see the protocol) to achieve the final OD600 = 0.5. (E) Gently mix the re-suspended cells in a Petri dish. (F) Use a needleless syringe to push the infiltration mixture into the abaxial side of the tobacco leaves. (G) About 48 hr after infiltration, the leaves are ready for confocal imaging. Excise leaf disks containing infected cells with a hole puncher around the infiltration region. Dashed circles indicate excised leave disks. (H) Use water to mount the leaf disks onto a standard slide for confocal imaging. The box diagrams in the shaded area describe the proteins injected into tobacco leaves for polarization assay. Protein sizes are not in scale. Please click here to view a larger version of this figure.
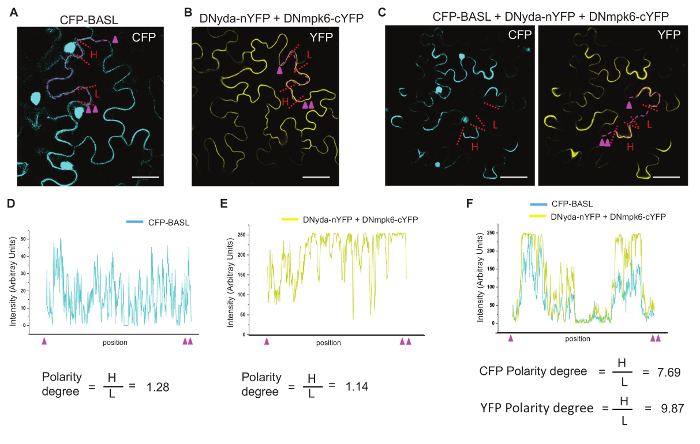
Figure 3. Display and Quantification of Polarity Formation. (A-C) Confocal images to show CFP and YFP florescence in tobacco epidermal cells. Cyan indicates the expression of CFP. Yellow indicates the expression of YFP. (A) Overexpression of CFP-BASL alone. (B) Co-expression of DNyda-nYFP and DNmpk6-cYFP. The YFP expression is complemented when two proteins interact. (C) Co-expression of CFP-BASL, DNyda-nYFP and DNmpk6-cYFP. Uneven distribution (polarity) is evident for both CFP and YFP. Scale bar = 50 µm in (A-C). (D-F) Graphically plotting the CFP/YFP fluorescence intensity along the dashed lines (Magenta) in (A-C). Magenta triangles mark the start and end point of the regions used for intensity plotting. Quantifications of the polarity degree in tobacco cells are presented underneath. The values of florescence intensity (H for high and L for low) are measured and collected by Fiji from the regions traced with red dashed lines in (A-C). For each sample, the values from 20 cells are averaged for significance test. Please click here to view a larger version of this figure.
