All procedures were performed in accordance with European Community standards for the care and use of laboratory animals, with the approval of the local ethics committee for animal experimentation (Ile de France-Paris-Committee, Authorization 4270).
1. Anesthesia
- Infuse a lethal dose of pentobarbital (up to 1 mg/10 g body weight) intraperitoneally (27-gauge needle and 1-ml syringe) into adult mice before surgery.
2. Vessel Perfusion
NOTE: There is no need to apply vet ointment to the eyes during vessel perfusion. This procedure is rapid (5-10 minutes) and ends in the death of the animal. Confirm the lack of response with a toe pinch.
- Using iris scissors, make an incision, about 4 cm long, into the abdominal wall and peritoneum, just beneath the rib cage.
- Make a small incision (a few millimeters long) in the diaphragm and then continue the incision of the diaphragm along the entire length of the rib cage to expose the pleural cavity.
- Lift the sternum away and clamp the tip of the sternum with the hemostat; place the hemostat on the neck. Carefully trim the adipose tissue connecting the sternum to the heart.
- Pass the 15-gauge perfusion needle through the left ventricle into the apex of the heart.
- Finally, use scissors to cut one of the liver lobes to create an outlet.
NOTE: An alternative outlet can be created by using iris scissors to create an incision to the right atrium. - Perfuse the animal with 25 to 50 ml of phosphate-buffered saline (PBS) with a pump operating at a rate of 2.5 ml/min. The liver should blanch as the blood is replaced with PBS.
- After approximately five minutes, once the fluid from the liver is completely clear, stop the perfusion.
- If immunostaining or regular staining is planned, perfuse the animal with 50 ml of paraformaldehyde (PFA; 4% in PBS) for 15 min.
NOTE: Caution, PFA fumes are toxic. Perfusion of the animal with PFA should be carried out in a ventilated fume hood.
3. Isolation of the Brain and the Circle of Willis
- Isolation of the brain
- Remove the head with a pair of surgical scissors.
- Make a midline incision with iris scissors, along the skin from the neck to the nose.
- Trim off the skin to expose the skull and remove any residual muscles and adipose tissues with iris scissors.
- Place the sharp end of the iris scissors into the foramen magnum on one side and carefully slide them along the inner surface of the skull to the external auditory meatus (also known as the ear canal).
- Reproduce the incision described in 3.1.4 on the contralateral side and make a midline cut along the inner surface of the inter-parietal bone to the start of the sagittal suture.
- Plant the iris scissors in the frontal bone, right between the eyes, in the sagittal suture and then open them to split the skull in two.
- Lift out the brain, grabbing the olfactory bulbs and using the iris scissors to cut off the nerve connections on its ventral surface.
- Remove the brain and place it in a 60-mm Petri dish containing ice-cold PBS for CoW isolation. Completely immerse the brain in the PBS. If the brain was fixed with 4% PFA (for subsequent sectioning and immunostaining or regular staining), keep it in a bath of 4% PFA at 4 °C for 24 hr.
- Isolation of the circle of Willis
NOTE: A dissecting microscope is required for CoW isolation. The brain should be kept at 4 °C throughout the entire procedure.- Put the brain upside down (i.e., on its dorsal surface) to visualize the CoW.
- Use a small forceps to grab the anterior cerebral arteries (ACA) at the base of the olfactory lobes (a, Figure 1) and exert pressure to dissociate them from the vessel continuum. Use the same procedure to cut the middle cerebral arteries (MCA) in b (Figure 1).
- Use the sharp ends of the forceps to lift and remove the major arteries forming the CoW from the cortex.
- Lift up the start of the posterior communicating arteries (PCA) to disconnect them from the brain, by gripping the middle cerebral arteries (MCA) with the forceps. Pick up the anterior arteries (ACA and MCA) and pull them gently over the optic chiasm in an anterior-dorsal direction. To prevent disruption of the CoW, interrupt the the procedure to deal with the other arteries.
- Repeat steps 3.2.2 and 3.2.3 for the superior and posterior cerebellar arteries (SCA)/(PCA) (c, Figure 1) and for the basilar artery (BA) (d, Figure 1), pulling them in a dorsal-anterior direction. Stop at the end of the procedure described in 3.2.4.
- Remove the entire CoW by pulling gently with the forceps. Place the CoW in a 60-mm Petri dish filled with ice-cold PBS and remove any remaining attached brain tissue with two forceps, holding the CoW in place with small pins.
- Keep the harvested CoW at -80 °C for subsequent processing for RNA purification (RNA extraction yields large amounts of RNA — approximately 500 ng) or protein extraction.
Note: The CoW can be maintained ex vivo for 24 hr, by adapting the organ bath system developed for isolated mouse olfactory artery. 7
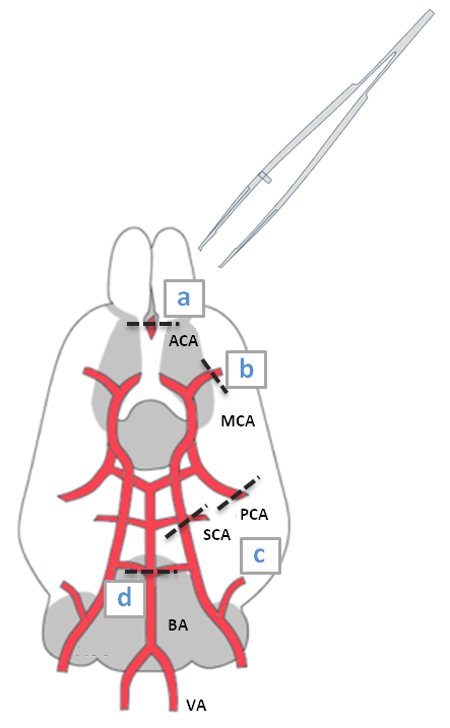
Figure 1: Schematic Diagram of a Ventral View of the Mouse Brain Highlighting the CoW. The CoW is formed from the two internal carotid arteries (MCA), which are derived from the two anterior cerebral arteries (ACA); the basilar artery (BA) branches into the posterior (PCA) and superior (SCA) cerebral arteries, and two vertebral arteries (VA).
The PBS-perfused mouse is killed and the CoW is isolated as described in section 3.2 of the protocol. When the dissection is performed correctly, the CoW should come out in one piece and should be slightly transparent due to the absence of residual blood in the vasculature.
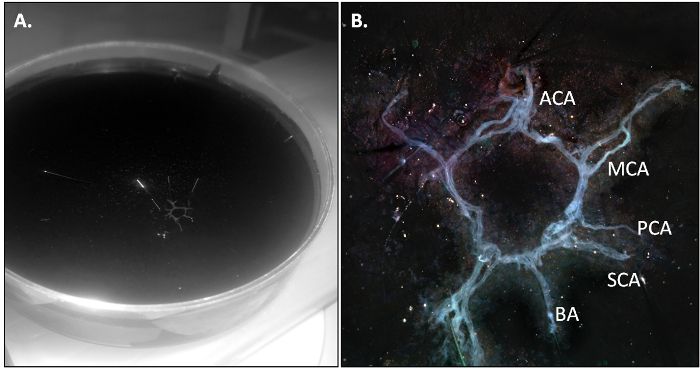
Figure 2: The Mouse CoW after Isolation. (A) Overview of the CoW in a 10-cm Petri dish. (B) Details of the various branches of the CoW. MCA for middle cerebral arteries, ACA for anterior cerebral arteries, BA for basilar artery, PCA for posterior cerebral arteries and SCA for superior cerebral arteries. Please click here to view a larger version of this figure.
The purity of the CoW preparation can be checked by ensuring that specific vascular genes are strongly expressed whereas the expression of neuronal genes is undetectable. More specifically, the cleaned CoW should specifically express vascular smooth muscle cell markers, such as smooth muscle actin (SMA), smooth muscle-myosin heavy chain (SM-MHC) and smoothelin. These markers are barely expressed in other parts of the brain. An opposite pattern of expression should be obtained for neuronal genes, with neuronal markers (Mog, Map2) only barely detectable in the CoW.
Levels of mRNA can be assessed by RT-qPCR with normalization relative to a reference gene transcript (here, that of GAPDH). Typical results are shown in Figure 3A and B.
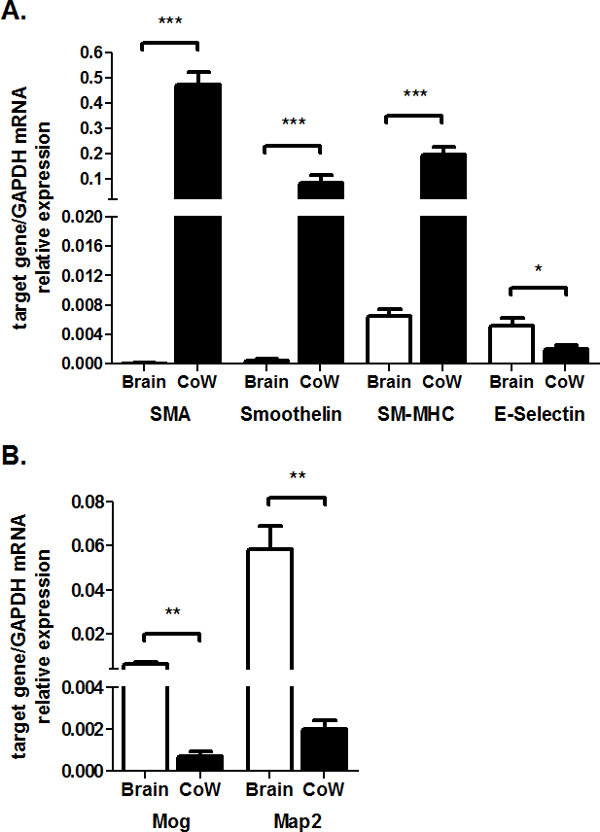
Figure 3: Expression of Neuronal and Contractile Genes in the Mouse Circle of Willis and Brain. The RT-qPCR results were normalized against those for a reference gene (GAPDH). They are expressed as the mean ± SD of 6 to 12 independent experiments; Comparison of the CoW with the brain: n.s, not significant, *, P < 0.05, **, P < 0.005, and ***, P < 0.001. (A) Expression of contractile genes: SMA (smooth muscle actin), smoothelin, SM-MHC (smooth muscle-myosin heavy chain 11) and E-selectin. (B) Expression of neuronal genes: Mog (myelin oligodendrocyte glycoprotein) and Map2 (microtubule associated protein 2).
The expression pattern of the endothelial marker E-selectin in brain lacking the CoW is very similar to that obtained for CoW samples, possibly reflecting the existence of a brain capillary network. Table 1 lists the mean Cp numbers for each transcript.
| A. | |||||||||||||||
| Mean Cp | GAPDH | SMA | Smoothelin | SM-MHC | E-Sel | ||||||||||
| Mean | SD | N | Mean | SD | N | Mean | SD | N | Mean | SD | N | Mean | SD | N | |
| CoW | 20.86 | 0.93 | 12 | 22.02 | 0.67 | 12 | 25.29 | 1.89 | 12 | 23.37 | 0.80 | 12 | 30.56 | 1.66 | 12 |
| Brain | 18.71 | 0.72 | 6 | 33.91 | 2.06 | 6 | 31.08 | 3.17 | 6 | 26.05 | 0.73 | 6 | 28.45 | 0.63 | 6 |
| B. | |||||||||||||||
| Mean Cp | GAPDH | Mog | Map2 | ||||||||||||
| Mean | SD | N | Mean | SD | N | Mean | SD | N | |||||||
| CoW | 21.04 | 0.41 | 6 | 32.13 | 1.63 | 6 | 30.21 | 0.85 | 6 | ||||||
| Brain | 19.09 | 0.49 | 6 | 26.43 | 0.88 | 6 | 23.30 | 0.99 | 6 | ||||||
Table 1: Mean Crossing Point for the Expression of Neuronal and Contractile Genes in the Mouse Circle of Willis and Brain. Calculations are based on comparison of the precise cycle determined by crossing points (Cp) at a constant level of fluorescence. The crossing point is the cycle number corresponding to the maximum second derivative of the amplification curve. Higher Cp values are associated with lower levels of expression. For a given gene, expression is considered to be undetectable if the Cp exceeds 35. (A) Cp of the reference gene: GAPDH; and of contractile genes: SMA, smoothelin, SM-MHC and E-selectin. (B) Cp of the reference gene: GAPDH; and neuronal genes: Mog and Map2.
