The described imaging protocol can be used to analyze the behavior of aggregation-prone prion-like proteins in the amoeba D. discoideum during heat stress. Figure 1 shows the distribution of GFP-tagged polyglutamine protein Q103 in cells under normal growth conditions (Figure 1A), after 2 hours of heat stress at 30 °C (Figure 1B) and after 6 hours of stress recovery and growth at normal growth conditions (Figure 1C). Upon temperature increase from 23 °C to 30 °C, the Q103-GFP marker coalesces from a diffuse distribution to punctate structures. Upon stress release and growth at normal temperature, these structures dissolve. The redistribution of Q103-GFP and formation of heat stress-induced cytosolic foci can be quantified using image analysis. Figure 2A shows the analysis of individual FOVs using Fiji and the plugin "Cell counter". Figure 2B shows the quantification of the heat stress response. This shows that the cytosolic Q103-GFP foci increase in number with continuing heat stress. During heat stress, the protein quality control (PQC) system is overwhelmed, thus aggregation-prone proteins such as the prion-like protein Q103 cannot be maintained in a soluble state and increasingly aggregate in cytosolic foci. The cells also show a characteristic rounding, as depicted in Figure 1B. The reduction of cytosolic foci after stress removal suggests that the aggregated proteins are dissolved.
The imaging quality is sensitive to the initial cell density. Figure 3 shows an example of cells after stress recovery, comparing appropriate initial cell density (Figure 3A) and high initial cell density (Figure 3B). If the initial cell density was too high, cells started to form streams as depicted in Figure 3b. This which imposes difficulties on the subsequent quantification as the cells have moved out of the focal plane.
The described method can be used to monitor temperature-induced changes as described above. It can also be used to reduce photo-toxicity when imaging at physiological growth temperature. Figure 4 shows a comparison of cell imaged at 23 °C in a cooling chamber (Figure 4A) and cells imaged without temperature control in an air conditioned room, set to 25 °C (Figure 4B). Upon constant exposure to the excitation light, cells round up when not imaged under temperature-controlled conditions. This change in cell shape is indicating stress.
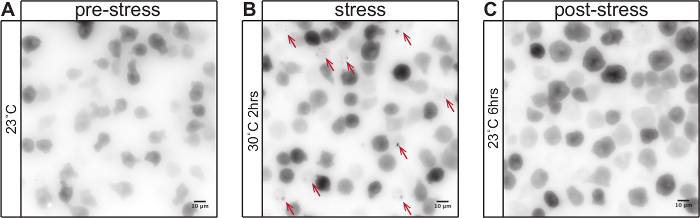
Figure 1: Aggregation-prone prion-like proteins change their cellular distribution during heat stress. GFP-tagged polyglutamine-rich huntingtin exon 1 (Q103) is constitutively expressed in AX2-214 D. discoideum cells (GFP signal depicted in inverted gray scale lookup table). (A) Under normal growth conditions at 23 °C, Q103-GFP is diffusely distributed in the cytoplasm. (B) After 2 hr of heat stress at 30 °C, Q103-GFP coalesces into cytosolic foci (red arrows). The cells show a characteristic shape change and round up. (C) Upon stress release and growth at 23 °C, the cytosolic foci are dissolved. After 6 hours, no cytosolic foci can be observed. Scale bar = 10 µm. Please click here to view a larger version of this figure.
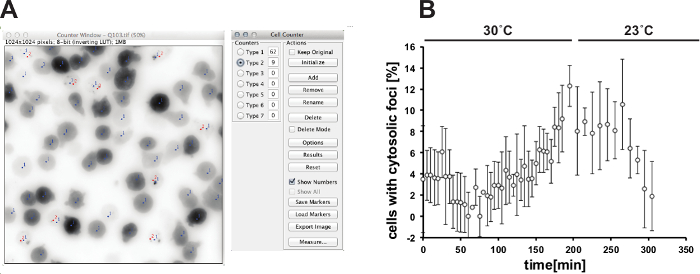
Figure 2: Quantification of heat-induced cytosolic foci formation. (A) Analysis of cells using the free image-processing package Fiji and the plugin "Cell Counter". The total amount of cells (type 1, blue counter markers) is determined in the bright field image. The markers are loaded in the GFP-channel and the number of cells with cytosolic foci (type 2, red counter markers) is determined. (B) Quantification of foci after 3 hr of heat stress at 30 °C and 3 hours of stress recovery at 23 °C (n = 4 fields of view, error bar = SD). Please click here to view a larger version of this figure.
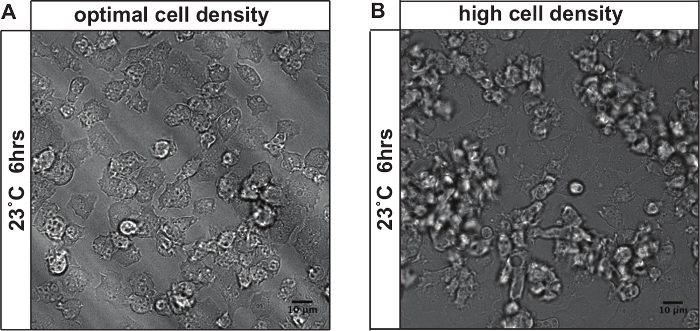
Figure 3: Effect of cell density on long-term time-lapse imaging. (A) Cells imaged at initial cell densities <105 cells/ml. After 3 hr of heat stress at 30 °C and 6 hr of stress recovery at 23 °C, cells still remain in the vegetative cycle and divide. Only few cells exit the focal plane. (B) Cells imaged at initial cell densities >106 cells/ml. After 9 hr of imaging (settings as in A), cells have entered the developmental cycle and started streaming and aggregating. Cells in the aggregates cannot be clearly distinguished. A large portion of cells exits the focal plane and cannot be analyzed. Scale bar = 10 µm. Please click here to view a larger version of this figure.
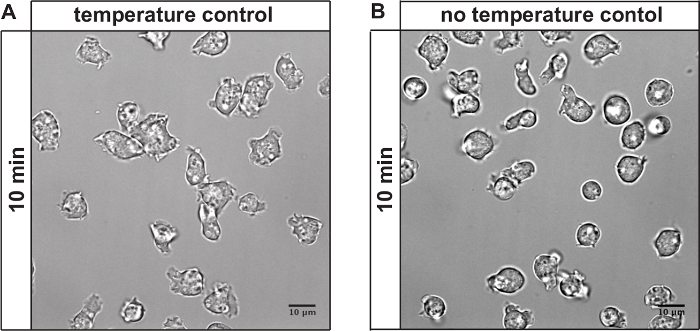
Figure 4: Effect of temperature control on phototoxic effects. (A) Cells imaged for 10 min with temperature control at 23 °C: 6 fields of view, z-stack (spacing 0.5 µM, sample thickness 8 µM), time lapse (total time 10 min, lapse 1 min), GFP channel (0.07 msec exposure time, 10% laser intensity), RFP channel (0.1 msec exposure time, 10% laser intensity). Cells show no signs of the characteristic morphological changes associated with phototoxic stress. (B) Cells imaged for 10 min without temperature control (same conditions as in A). Cells exhibit signs of phototoxic induced rounding, indicating stress. Scale bar = 10 µm. The cells were imaged on a system based on a microscope with a 100x/1.4 numerical aperture (N.A.) objective. The images were collected with a CCD camera as 1,024 x 1,024 pixel files using 1 x 1 binning. Please click here to view a larger version of this figure.
