While the interaction between Drp1 and Mff has been demonstrated to be important for mitochondrial fission, this interaction has been difficult to recapitulate in vitro. Our goal was to better emulate the cellular environment wherein Drp1 and Mff interact. To this end, liposomes containing limiting concentrations of NTA(Ni2+) headgroups were prepared by rehydrating a lipid film as described above. The lipid solution initially consists of unilamellar and multilamellar vesicles of heterogeneous diameters as evidenced by the opacity of the solution (Figure 1a). This opacity is reduced by freeze-thawing (Figure 1b), which reduces the prevalence of multilamellar vesicles. The liposome diameters are further homogenized by extrusion through a polycarbonate filter, which results in a clear solution (Figure 1c).
In previous studies, we found that Drp1 was able to assemble on Mff-decorated scaffold liposomes, and membrane tubulation was observed when flexible membranes were employed2. Building on these findings, we utilized a new template composed of PC, PE, Ni, and CL (called Enriched Scaffold Liposomes or ESL) to promote ordered assembly of a polymeric Drp1-Mff complex capable of inducing membrane deformation. Specifically, increased NTA(Ni2+) and cardiolipin lipids were utilized (10 mol% and 15 mol% respectively) for this application. Then, GFP or Mff was tethered to ESL templates in the presence and absence of Drp1 (Figure 2), and the ability of Drp1 to remodel membranes was qualitatively assessed. In the absence of Drp1, neither Mff nor GFP resulted in membrane deformation (Figure 3a, b), and similarly in the case of GFP-decorated ESL, only featureless liposomes were observed (Figure 3c). However, when Drp1 was added to Mff-decorated ESL templates, remodeling of the liposomes was evident (Figure 3d).
While macromolecular complex formation clearly demonstrates an interaction between Drp1 and Mff, this qualitative analysis alone is incapable of determining the functional effects of such an interaction. Therefore, we utilized a malachite green phosphate generation assay18 to assess alterations in the catalytic activity of Drp1 in response to interaction with Mff. As described previously2, we initially utilized a simple scaffold liposome (SL; 3.3 mol% DGS-NTA(Ni2+), 96.7 mol% DOPC) to investigate the effect of Mff alone on Drp1 structure and function. Nonspecific interaction of Drp1 with NTA(Ni2+) has previously been described20, so SL was initially designed to contain low concentrations of NTA(Ni2+) to avoid nonspecific activity stimulation of Drp1. With the larger amounts of NTA(Ni2+) in ESL, the use of His-tagged GFP as a control was found to be critical to shield the Ni2+ and prevent non-specific Drp1 interactions. After decoration of SL liposomes by Mff or GFP (as illustrated in Figure 2), the extent of self-assembly can be assessed by measuring the GTPase activity of Drp1. In the absence of liposomes, Drp1 has a relatively low basal GTPase activity, which is slightly enhanced by addition of SL. Decoration of these scaffold liposomes with Mff enhanced GTPase activity (Figure 4a, 1.8 fold). Conversely, when the exposed NTA(Ni2+) headgroups were blocked with His-tagged GFP, this augmented GTPase activity was ablated. We also tested the role of Fis1, an OMM protein that has been suggested to have a role in mitochondrial fission21,22, though this has been challenged in recent studies7,23. Tethering of Fis1 lacking its transmembrane domain to SL also failed to elicit a stimulation of Drp1's GTPase activity (Figure 4a).
We then utilized a slightly more complex lipid scaffold containing a small amount of cardiolipin (SL/CL: SL with 10 mol% cardiolipin replacing DOPC) to determine the role of this mitochondrial lipid in the interaction of Drp1 and Mff. This moderate concentration of cardiolipin was specifically chosen to limit the stimulation of Drp1 by cardiolipin as described previously10. Similar to SL, addition of SL/CL to Drp1 resulted in a slight stimulation of GTPase activity that was reversed by tethering His-tagged Fis1 or GFP to the liposomes. A synergy between Mff and cardiolipin was observed as the GTPase activity of Drp1 was stimulated 2.6 fold when it was incubated with Mff-decorated SL/CL (Figure 4b).
Membrane fluidity and the ability of Drp1 to remodel lipid bilayers have been proposed to enhance its GTPase activity. Therefore, we sought to examine the effect of membrane fluidity/flexibility using a flexible scaffold liposome. This was achieved by replacing 35 mol% of DOPC in SL/CL with DOPE (SL/PE/CL), which has previously been shown to allow for Drp1-mediated membrane remodeling10. Addition of undecorated SL/PE/CL scaffold liposomes to Drp1 slightly enhances Drp1 GTPase activity, and decoration of these liposomes with GFP eliminates this effect. When SL/PE/CL templates were decorated with Mff, Drp1 activity was enhanced (Figure 4c, 2.4 fold). As we have previously shown, the ability of Drp1 to remodel liposomes into lipid tubules was enhanced by the addition of PE to the scaffold liposomes. Interestingly, this improved tubulation leading to the formation of a helical Drp1 polymer did not result in any greater stimulation when compared to liposomes that Drp1 was unable to remodel2.
Using these adaptable lipid templates, Mff and Drp1 were found to interact in a more native environment in vitro. This technique has enabled us to control the relative abundance of Drp1, Mff (through NTA(Ni2+) concentration), and specific lipids (cardiolipin and PE specifically) that appeared to regulate the assembly of this macromolecular complex. As we have demonstrated, this method can be utilized to visualize the membrane remodeling of Mff-recruited Drp1 by electron microscopy, and to determine the effects of Drp1 assembly on its catalytic activity using GTPase activity assay.
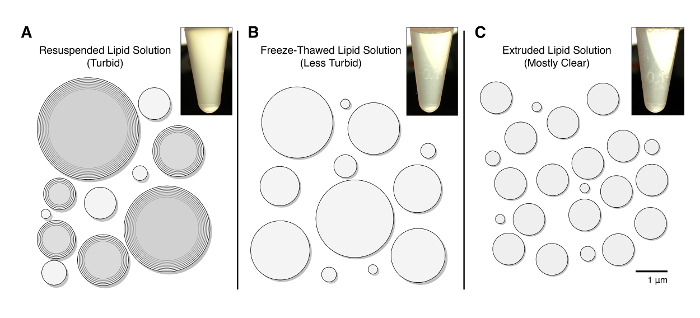
Figure 1: Lipid Preparation Schematic. (a) Upon resuspension, liposomes of diverse sizes form and consist of unilamellar and multilamellar vesicles, which results in an opaque solution (inset). (b) Freeze-thawing the solution results in a more unilamellar population of liposomes, which are still heterogeneous in diameter. Freeze-thawing clarifies the solution (inset). (c) Extrusion of the lipid solution homogenizes the liposome diameter (1.0 µm in this example), and results in a clear solution (inset). Please click here to view a larger version of this figure.
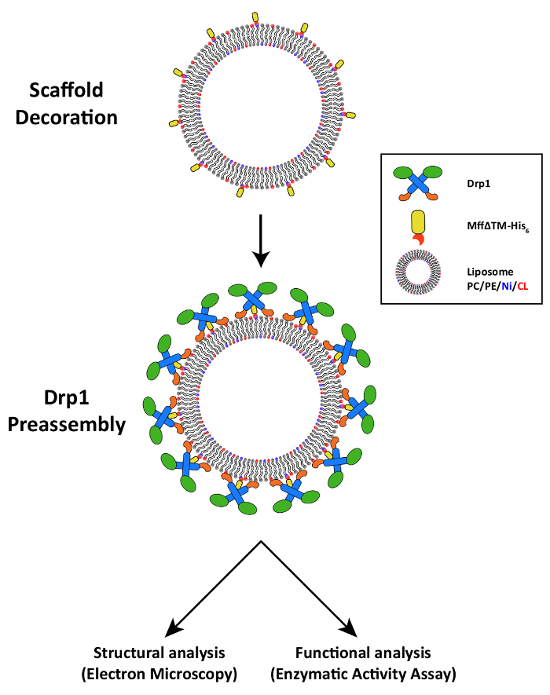
Figure 2: Methods to Assess Protein assembly. A schematic depicting partner protein assembly on scaffold liposomes is presented. His-tagged partner proteins or GFP are incubated with scaffold liposomes, and then Drp1 is incubated with decorated or undecorated liposomes. These Drp1-preassembled liposomes can then be analyzed by structural methods (electron microscopy) and functional assays (GTPase assay). Please click here to view a larger version of this figure.
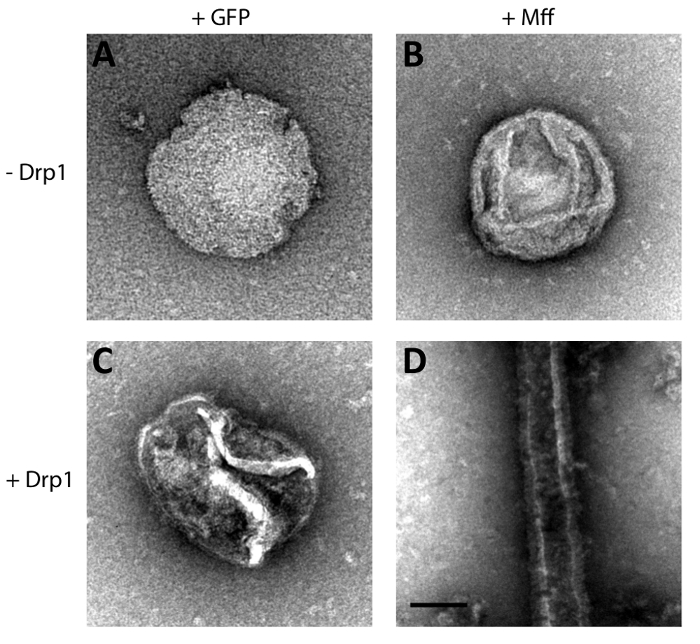
Figure 3: Structural Assessment of Drp1 Recruitment. Negative stain transmission micrographs of GFP or Mff decorated liposomes alone (A, B, respectively) or incubated with Drp1 (C, D, respectively). Scale bar = 100 nm. Please click here to view a larger version of this figure.
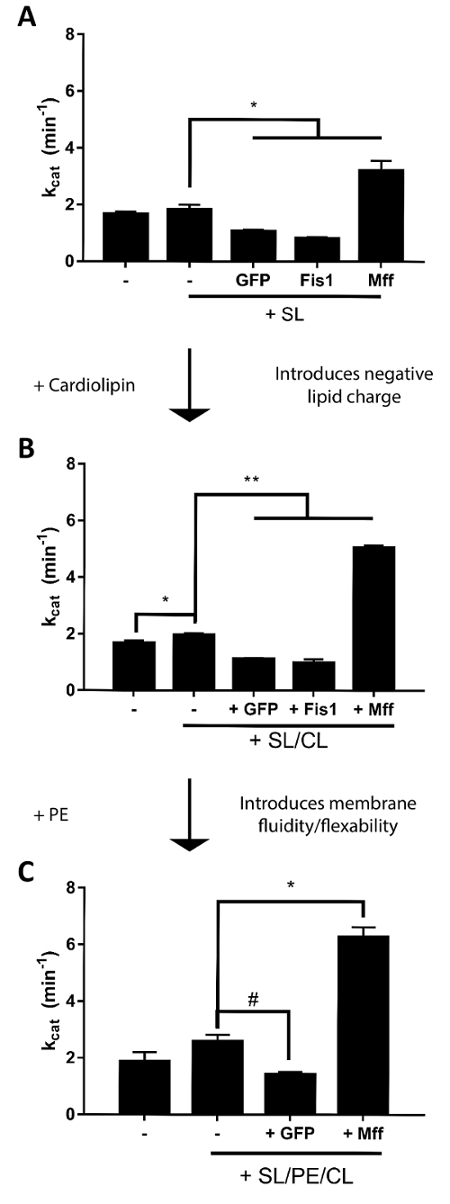
Figure 4: Scaffold Liposome Enzymatic Assay. (A – C) The generation of phosphate over time was measured (inset), and the kcat was determined. This method was applied to SL-tethered proteins (A), SL/CL-tethered proteins (B), or SL/PE/CL-tethered proteins (C). #: p <0.05, *: p <0.0001, **: p <0.000001 as determined by unpaired student's T-test. All error bars represent standard deviation from 3 independent samples. Please click here to view a larger version of this figure.
