Please note that this protocol was successfully used with a number of kidney GFP transgenic lines, including ET(krt8:EGFP)sqet11-9, ET(krt8:EGFP)sqet33-d10, and Tg(atp1a1a.4:GFP). The example results shown here were obtained using the ET(krt8:EGFP)sqet11-9 line.
Figure 1 shows the example photoablation protocol. The average GFP intensity is monitored inside the region of interest (Figure 1A, and Movie 1). An example average intensity trace is shown in Figure 1B.
As shown in Figure 2, after exposure to 405 nm laser light, GFP fluorescence continues to disappear one cell at a time until, at 220 min, the entire ablated segment loses 100% of its GFP positivity. This loss of GFP fluorescence is indeed due to cell death and not, for example, the downregulation of GFP expression. This is evidenced by the appearance of red fluorescent propidium iodide-positive nuclei in the cells that lose GFP positivity.
The extent of cell death can be assessed by measuring average GFP fluorescence in the ablated segment and comparing it to the mean GFP intensity immediately anterior and posterior to the ablated segment. Figure 3 shows the relationship between the extent of laser exposure (measured by the initial amount of photobleaching) and the extent of cell death in the exposed segment. It shows that, by varying the initial exposure, it is possible to "dial" the injury and the response of the injured epithelium. With 10-20% of initial GFP photobleaching, virtually no reduction of GFP positivity is observed at 5 h post-injury. 50 and 60% photobleaching leads to a complete disappearance of GFP at 5 h, while 30 and 40% bleaching leads to intermediate results, with 50% estimated death observed at about 35% photobleaching. These results are supported by PI staining (Figure 3B). 20% photobleaching results in virtually no PI staining at 100 min post-ablation. 50% photobleaching leads to continuous PI positivity in the ablated epithelium after 60 min, while 30 and 40% photobleaching leads to intermediate PI incorporation. As can be seen from this data, this methodology allows for the induction of graded amounts of epithelial injury and for the study of the epithelial response to lethal and sub-lethal epithelial damage.
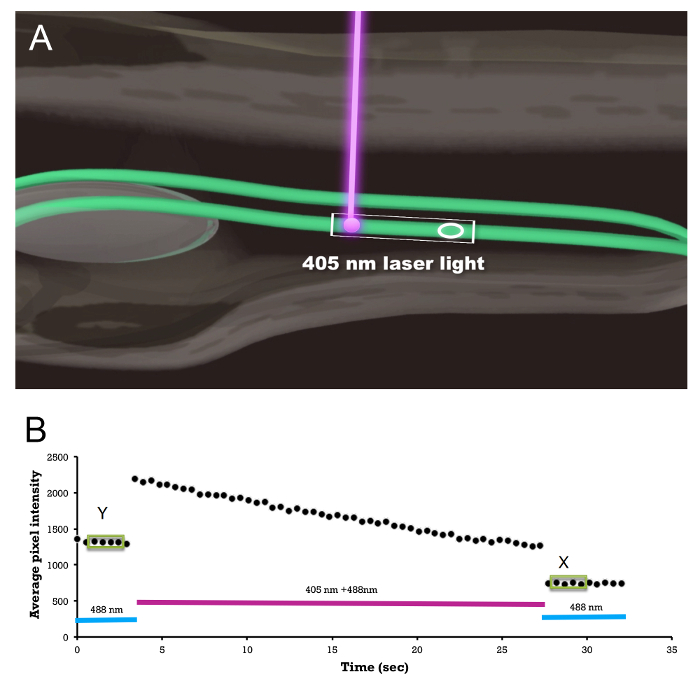
Figure 1: Photoablation Procedure. (A) Schematic showing embryo/larva orientation, laser exposure, and the region-of-interest (ellipse) measurement of the average fluorescence to monitor the amount of GFP photobleaching; see Movie 1. The rectangular box indicates the scan window. (B) An example of an actual region-of-interest average GFP intensity trace before, during, and after 405 nm laser exposure. Average GFP intensities on four consecutive measurements (shown inside green boxes) are shown before (Y) and after (X) the photoablation. The ratio X/Y is used as a measure of the total light exposure and is plotted on the X axis in Figure 3. Please click here to view a larger version of this figure.
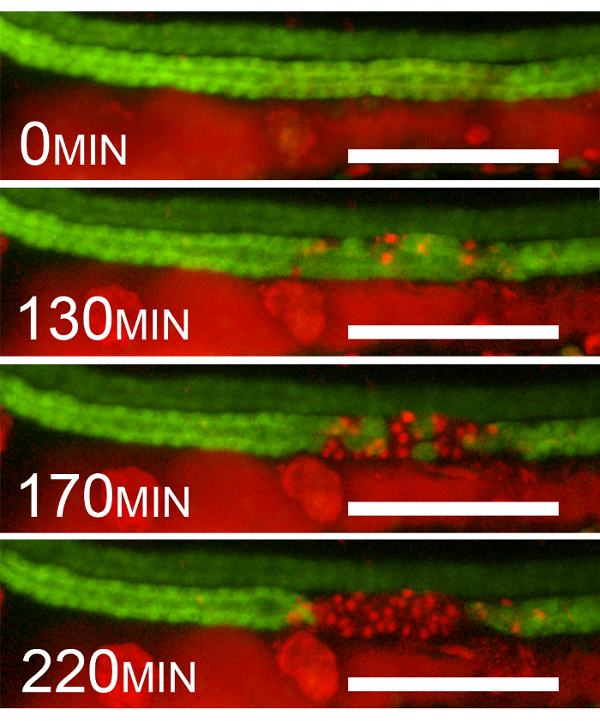
Figure 2: Epithelial Cell Death after 405 nm Laser Exposure. Example sequential frames showing the disappearance of GFP and the appearance of propidium iodide staining after photoablation (40% initial photobleaching). The frames are at 0, 130, 170, and 220 min after violet-light (405 nm) exposure. The disappearance of GFP coincides with the appearance of red fluorescent PI nuclear positivity. Please note that PrI fluorescence is detected by using the red channel. The red fluorescence seen outside the kidney is due to chromophores and the presence of PI in the gut lumen. Scale bar = 100 µm. Please click here to view a larger version of this figure.
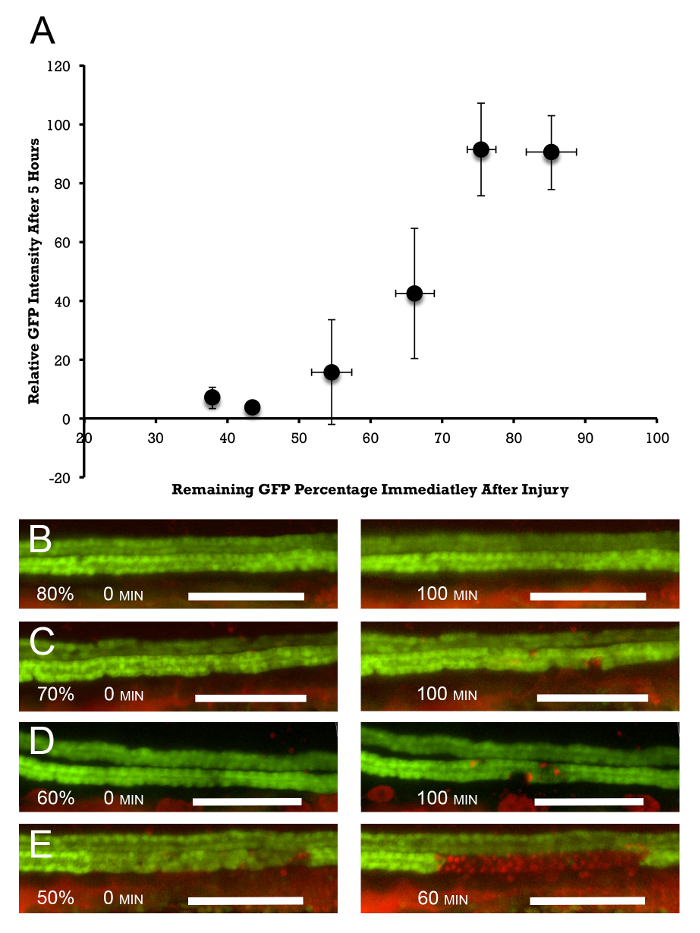
Figure 3: Dose Response of Epithelial Injury to the Amount of Photoablation. (A) Photoablation is measured by the percent of initial reduction in GFP fluorescence in the exposed segment. This is compared to the decline in average GFP fluorescence in the injured segment at 5 h post-injury. This measurement is normalized to the average amount of fluorescence upstream and downstream of the injured segment. Target doses were 10, 20, 30, 40, 50, and 60% of the initial photobleaching. The actual amounts are slightly larger, amounting to 14.8, 24.4, 33.8, 45.4, 56.5, and 62.1% photobleaching, which result in 85.2, 75.6, 66.2, 54.6, 43.5, and 37.9% remaining fluorescence, n = 2 – 6/target group. The error bars represent the standard deviation along the X (percent of remaining fluorescence immediately after photoablation) and Y (percent of remaining fluorescence 5 h after photoablation) axes. (B-E) PI staining parallels the overall disappearance of GFP positivity. Virtually no PI staining is observed at 20% (80% remaining fluorescence) photobleaching at 100 min post-laser exposure (B, right panel). (C and D) 30 and 40% photobleaching (70 and 60% remaining fluorescence) show increasing numbers of PrI-positive cells (right panels), and (E) at 50% photobleaching, virtually 100% PI positivity is observed at 60 min post-injury (right panel). The left panels in (B-E) show the initial amount of photobleaching. Scale bar = 100 µm. Please click here to view a larger version of this figure.
| Transgenic | Expression Pattern | References |
| Tg(wt1b:GFP) | Glomerulus, some PT | [11] |
| Tg(atp1a1a.4:GFP) | Distal to glomerulus | [12] |
| Tg(cdh17:GFP) | Distal to glomerulus | [9,10] |
| Tg(ret1:GFP) | Late DT, PD | [13] |
| Tg(enpep:GFP) | Distal to glomerulus | [14] |
| Tg(cd41:GFP) | Multiciliated cells | [15] |
| ET(krt8:EGFP)sqet11-9 | Straight PT, early DT | [16,17] |
| ET(krt8:EGFP)sqet33-d10 | Convoluted PT | [16,17] |
| PT = Proximal Tubule | ||
| DT = Distal Tubule | ||
| PD – Pronephric Duct |
Table 1: Examples of Kidney GFP Zebrafish Lines. Some representative zebrafish lines are listed here, indicating which segment of the pronephric kidney is labeled in a particular transgenic line.
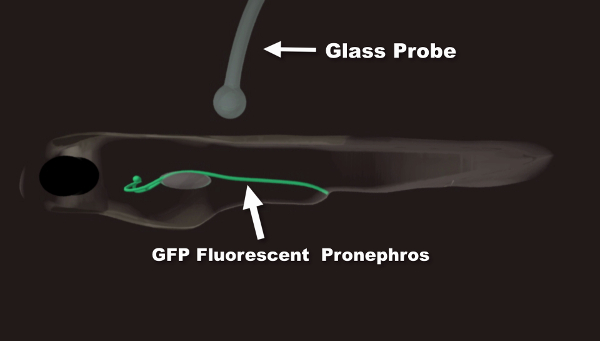
Movie 1: Animation Summary of the Photoablation Procedure. In the first part of the movie, proper embryo/larva orientation is demonstrated. The two kidney branches are shown in green. A glass probe is used to orient the fish in agarose. This is done if a control, non-injured branch is desired. Angling the fish allows for laser exposure on only one branch of the pronephric kidney. In the second part of the movie, the laser ablation procedure is outlined. The ellipse indicates the region of interest within the segment undergoing ablation used to monitor the amount of initial photobleaching. The rectangular window allows for the precise adjustment of the size and position of the ablated segment. Please click here to view this video. (Right-click to download.)
