The olfactory glomerular map is formed by initial global targeting and subsequent glomerular segregation of OSN axons1,2. Glomerular segregation is regulated by the adhesive/repulsive axonal interactions mediated by axon-sorting molecules whose expression levels are determined by expressed OR molecules7. The axon-sorting molecules involved in glomerular segregation are expressed in a position-independent mosaic manner in the OB9. In this study, we selected the following genes: Kirrel2, Sema7A, OLPC, BIG-2, and PCDH17. Some of these axon-sorting molecules are expressed in an activity-dependent manner7,8,10.
The quadruple immunostaining method shown here allowed the visualization of the expression patterns of four molecules simultaneously on the same section. These axon-sorting molecules showed position-independent mosaic expressions, but their patterns were not identical (Figures 1A, B). The glomerular structure was defined by fluorescent signals of VGLUT2 (Figure 2A). Axon-sorting molecules (Kirrel2, Sema7A, OLPC) were differentially expressed in each glomerulus; however, VGLUT2, which was used as a glomerular marker, was uniformly expressed among glomeruli (Figures 2B, 2C).
A single glomerulus can be considered as a data unit that is represented by multiple variables. To analyze this multivariate expression data, principal component analysis (PCA) was performed. PCA defines a new coordinate system, so that the first coordinate has the greatest variance datasets and that each succeeding coordinate has the greatest residual variance. The expression datasets were subjected to PCA and plotted in the space of the first and second principal component (PC1 and PC2) (Figure 3A). The contribution ratio of each principal component indicates how much percentage each principal component represents in the total tendency of the variables. The contribution ratios of PC1, PC2, PC3, PC4, and PC5 were 33.4%, 28.2%, 19.8%, 17.3%, and 6.3%, respectively (Figure 3B). PCA also revealed the factor loadings in each principal component. The squared factor loadings indicate how much percentage of the variance in an original variable is explained by a factor. In PC1, the factor loadings of Sema7A, OLPC, and Kirrel2 were positively loaded, whereas those of BIG-2 and PCDH17 were not (Figure 3C). A previous study analyzed the expression of these molecules in the mutant mice deficient for the cyclic nucleotide-gated (CNG) channel gene, which is a component of the olfactory signal transduction11, and showed that the patterns of CNG-dependent changes in the expression levels resemble the factor loadings in PC19. These results suggested that the variety of expressions of these molecules was generated by CNG channel-mediated neural activity.
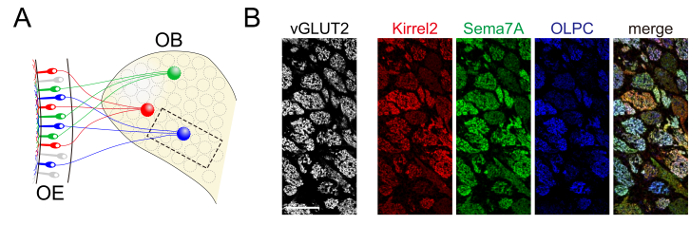
Figure 1: Quadruple Immunostaining of a Parasagittal OB Section. (A) Schematic diagram of the OE & OB. (B) A parasagittal OB section from a 2 week-old mouse immunostained with antibodies against v GLUT2 (white), Kirrel2 (red), Sema7A (green), and OLPC (blue). The merged image of Kirrel2 (red), Sema7A (green) and OLPC (blue) was shown at right. Scale bars = 100 µm. Please click here to view a larger version of this figure.
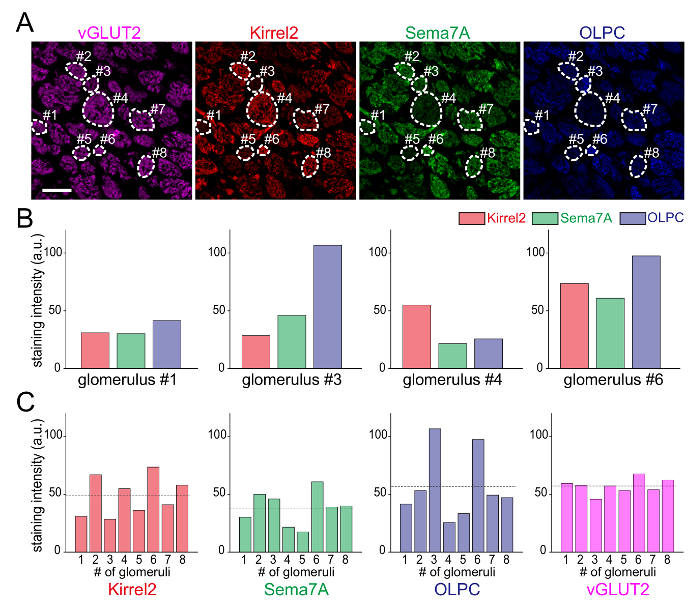
Figure 2: Expression Analysis of Axon-sorting Molecules. (A) A parasagittal olfactory bulb (OB) section immunostained with antibodies against VGLUT2 (purple), Kirrel2 (red), Sema7A (green), and OLPC (blue). Each glomerulus is defined by fluorescent signals of VGLUT2 (pre-synaptic marker). Glomerular structures are surrounded by dashed lines. (B) Expression levels of Kirrel2, Sema7A, and OLPC in glomeruli #1, #3, #4, and #6. Staining intensities of axon-sorting molecules were measured in each glomerulus. (C) Expression levels of the axon-sorting molecules and VGLUT2 compared among the glomeruli. Signal intensities of these molecules were measured in each glomerulus. Grey bars on the graphs indicate the average of expression level. Scale bar = 100 µm. Please click here to view a larger version of this figure.
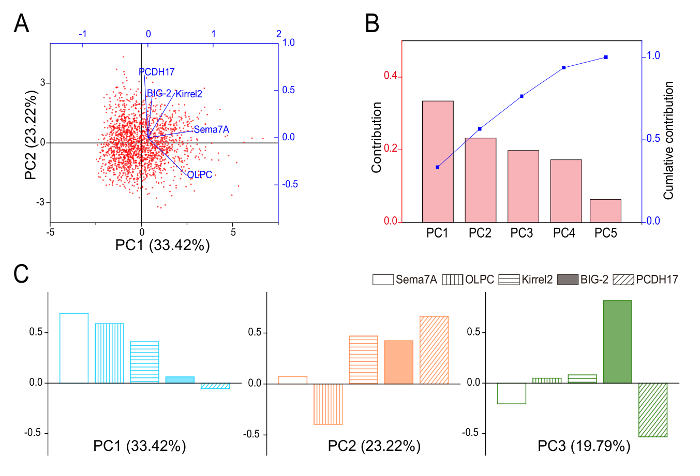
Figure 3: PCA of Expression Data of Axon-sorting Molecules. (A) Principal component analysis (PCA) score biplots (PC1 Vs. PC2) of the expression dataset of five axon-sorting molecules (Kirrel2, Sema7A, OLPC, BIG-2, and PCDH17). A total of 1799 glomeruli were analyzed. (B) The contribution ratios and cumulative contribution ratios of all five principal components. The contribution ratios of PC1, PC2, PC3, PC4, and PC5 are 0.334, 0.282, 0.198, 0.173, and 0.063, respectively. (C) The factor loadings of Kirrel2, Sema7A, OLPC, BIG-2, and PCDH17 in PC1-3. Eigenvalues of PC1, PC2, and PC3 are 1.67, 1.16, and 0.99, respectively. Please click here to view a larger version of this figure.
