Before using BCPPs to study intracellular interaction, it is critical to compare the effects of BCPP vs CPP to validate the results obtained with BCPP. Consequently, to study whether the inclusion of biotin modifies the activity of the target sequence, we first analyzed the effect of TAT-Cx43266-283-B compared with TAT-Cx43266-283 on G166 GSCs morphology. To do so, we performed some immunofluorescence analyses of two cytoskeletal proteins, F-actin and α-tubulin after 24 h of treatment. Figure 1 shows that G166 GSCs in the presence of 50 µM TAT-Cx43266-283 or TAT-Cx43266-283-B acquire a more rounded shape compared to the elongated and expanded cellular prolongations shown in the controls (TAT or TAT-B). In fact, Figure 1b shows that actin filaments are mostly assembled as actin networks when the cells were treated with TAT-Cx43266-283 or TAT-Cx43266-283-B while they form more actin bundles in the control cells (treated with TAT or TAT-B)25. In contrast, α-tubulin distribution does not vary between the different conditions. These results showed that the presence of biotin did not modify the effect of the target sequence on the morphology of G166 GSCs. In previous studies20,21, we showed that TAT-Cx43266-283 reduced G166 GSCs proliferation. In this study, we investigated whether TAT-Cx43266-283-B exerts the same effects in the growth as TAT-Cx43266-283. To do so, we analyzed the G166 GSCs proliferation by MTT assay after 72 h of treatment. The MTT assay is a colorimetric assay for assessing cell metabolic activity. MTT is metabolized by NAD(P)H oxidoreductase enzymes in mitochondria reflecting the number of viable cells present. Figure 2 shows that the reduction in the G166 GSCs cell viability is not significantly different when cells were treated with 50 µM TAT-Cx43266-283 or 50 µM TAT-Cx43266-283-B. Indeed, both significantly diminished G166 GSCs proliferation as compared to the control, TAT or TAT-B.
Once we confirmed that the effect of our target sequence in G166 GSCs (TAT-Cx43266-283) was not modified by the inclusion of biotin at the C-terminus (TAT-Cx43266-283-B), we investigated the intracellular partners of this sequence following the protocol described in this study (Figure 3). Because caveolae have been involved in the mechanism of TAT internalization26, we analyzed the presence of caveolin-1 (Cav-1) in the pull-downs. Western blot analysis (Figure 4) showed that TAT-B and TAT-Cx43266-283-B interact with Cav-1. However, the ability of TAT-Cx43266-283-B to recruit c-Src, PTEN and CSK is stronger than that found with TAT-B. Focal adhesion kinase (FAK) is a substrate of c-Src that has not been shown to interact with Cx43. Indeed, FAK did not show any significant interaction with either TAT-B or TAT-Cx43266-283-B.
To confirm the interaction between TAT-Cx43266-283-B and c-Src, G166 GSCs were incubated with 50 µM TAT-Cx43266-283-B for 30 min and their localization was followed with fluorescent streptavidin by confocal microscopy (Figure 5). Our results showed that the intracellular distribution of TAT-Cx43266-283-B is close to the plasma membrane (shown by phosphatidylserine staining) and matches with that of c-Src. In fact, co-localization analyses revealed some points of co-localization (white) between TAT-Cx43266-283-B and c-Src in the merge image. Consequently, confocal microscopy studies confirm the results obtained with the BCPP pull-down protocol described in this study.
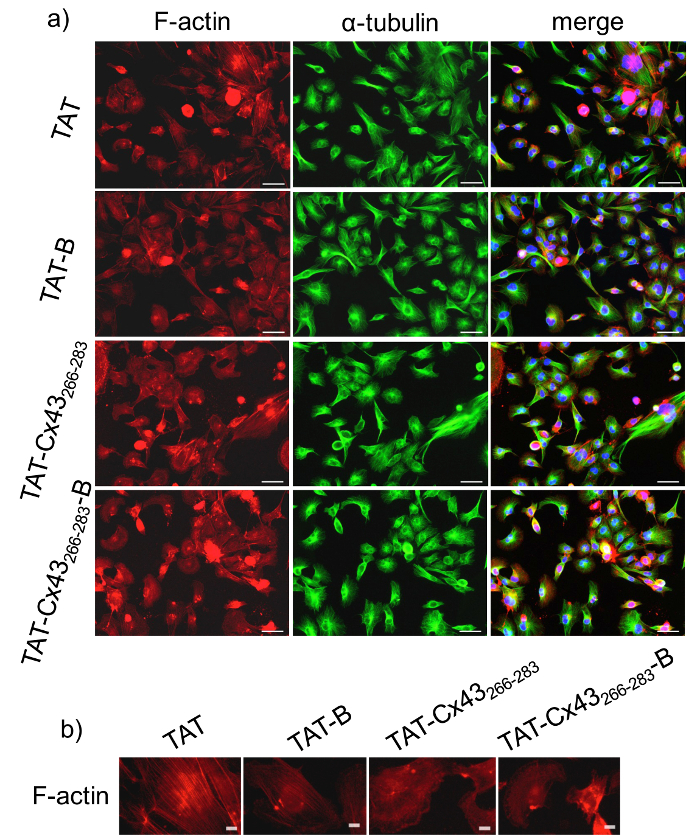
Figure 1: Effect of BCPP and CPP on GSC morphology.
G166 GSCs were plated at a low density (2 x 104 cells / cm2) and after 24 h they were incubated with 50 µM control CPP (TAT), control BCPP (TAT-B), treatment CPP (TAT-Cx43266-283) or treatment BCPP (TAT-Cx43266-283-B). a) F-actin (red), α-tubulin (green) and merged + DAPI immunostaining of the same field showing G166 GSCs morphology. Bars = 50 µm. b) F-actin immunostaining showing the different distribution of F-actin in G166 GSCs after incubation for 24 h with 50 µM control CPP (TAT) or control BCPP (TAT-B) as compared with 50 µM treatment CPP (TAT-Cx43266-283) or BCPP (TAT-Cx43266-283-B). Bars = 10 µm. Please click here to view a larger version of this figure.
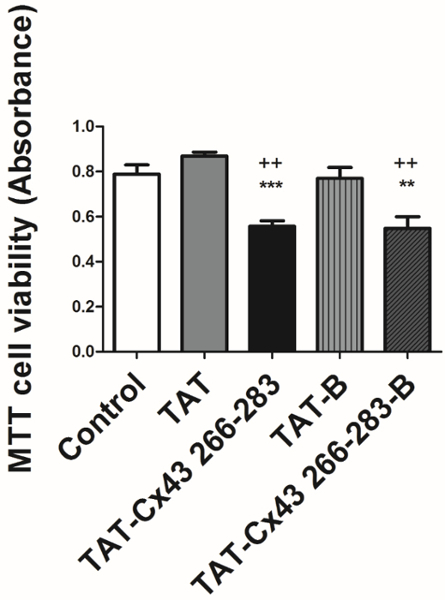
Figure 2: Effect of BCPP and CPP on GSC viability.
G166 GSCs were plated at 5500 cells/cm2 in 24-multiwell plates and incubated with 50 µM control peptides, CPP (TAT) or BCPP (TAT-B), or 50 µM treatment peptides, CPP (TAT-Cx43266-283) or BCPP (TAT-Cx43266-283-B). The cell viability was analyzed using a MTT assay after 72 h. The results are expressed as MTT absorbances and are the mean ± s.e.m. of at least 3 experiments (++ p˂0.01 vs control. ** p˂0.01, *** p˂0.001 vs TAT or TAT-B; one-way ANOVA withTukey post-test). Note that there are not significant differences between the effects of CPPs vs BCPPs. Please click here to view a larger version of this figure.
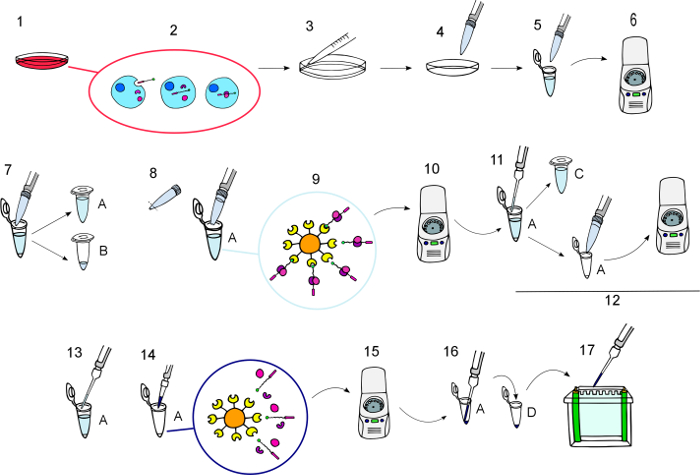
Figure 3: Protocol diagram.
Step by step graphical depiction of the procedure as described in the section "Protocol", from the incubation of the BCPPs until the eluted BCPPs and their interacting proteins were obtained.1) Incubate culture cells with BCPPs at the desired concentration for the required time. 2) During the incubation, the BCPPs are internalized and they interact with their intracellular partners. 3) Wash the cells three times on ice with ice-cold PBS. 4) Lyse the cells to extract proteins. 5) Transfer cell lysates to tubes. 6) Spin at 11000 x g for 10 min at 4 °C. 7) Transfer the supernatants to new tubes (A) and keep a small aliquot of the lysates to process as regular Western blot samples in tube (B). 8) Resuspend the NeutrAvidin Agarose beads and add 50 µL to each tube A using a cut pipette tip. 9) Incubate with gently shaking for 12 h at 4 °C to allow the NeutrAvidin agarose beads to interact with BCPPs and their partners. 10) Spin for 1 min at 3000 x g to pellet the beads with the biotinylated baits and their interacting proteins bound to them. 11) Transfer supernatants to new tubes (C) and keep them to use in case the pull-down need to be repeated. 12) Wash the pellet five times with fresh lysis buffer, resuspend by inversion, spin for 1 min at 3000 x g and discard the supernatant. 13) Remove all the supernatant carefully. 14) Add the desired volume of 4x Laemmli buffer and elute the proteins at 100 °C for 5 min. 15) Spin at 8200 x g for 30 s to pellet the beads. 16) Transfer the eluted proteins found in the supernatant with capillary tips to new tubes (D). 17) Load onto gels for Western blot analysis. Please click here to view a larger version of this figure.
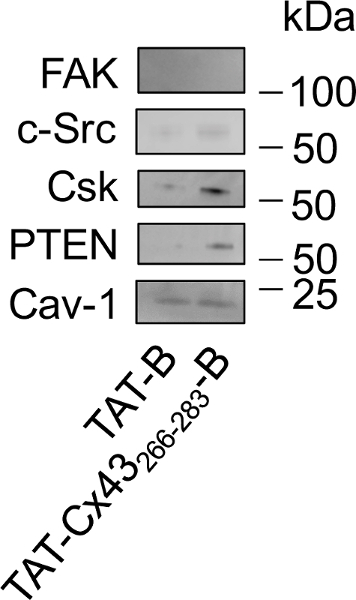
Figure 4: Study of the intracellular interactions of TAT-Cx43266-283-B in G166 GSCs by pull-down followed by Western blot.
G166 GSCs were incubated with 50 µM TAT-B or TAT-Cx43266-283-B. After 30 min the cells were lysed and TAT-B or TAT-Cx43266-283-B attached to their intracellular partners were pulled down with NeutrAvidin beads. The eluted proteins were loaded and analyzed by Western blot to study the levels of FAK, c-Src, CSK, PTEN and Cav-1. Note that Cav-1 interacts with both TAT-B and TAT-Cx43266-283-B, c-Src, PTEN and CSK interact preferentially with TAT-Cx43266-283-B and FAK did not show any interaction with either TAT-B or TAT-Cx43266-283-B. Please click here to view a larger version of this figure.
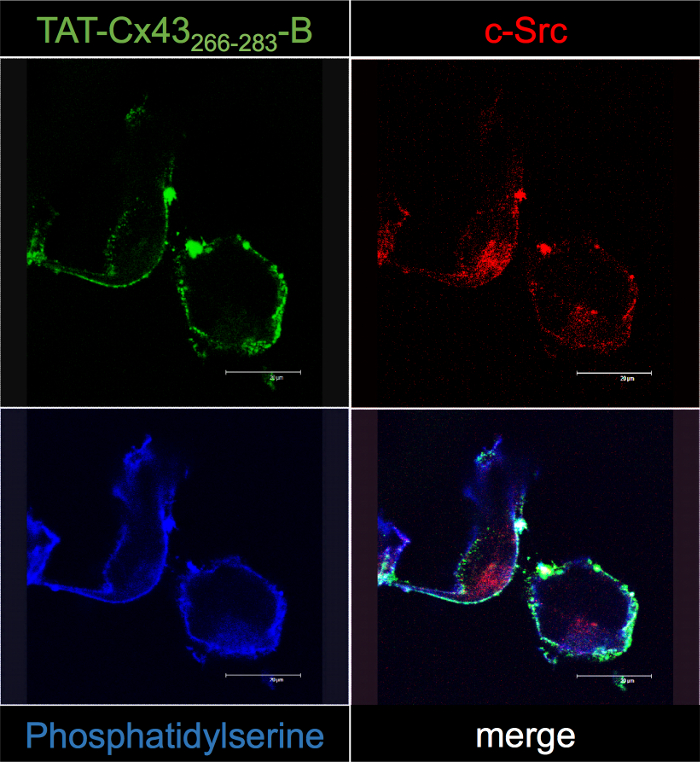
Figure 5: Confirmation of TAT-Cx43266-283-B intracellular interactions in G166 GSCs by confocal microscopy.
G166 GSCs were incubated with 50 µM TAT-Cx43266-283-B. After 30 min, cells were fixed and processed to localize TAT-Cx43266-283-B with Cy2-Streptavidin (green), c-Src by immunofluorescence (red) and phosphatidylserine with annexin V (blue). Note some points of co-localization (white) between TAT-Cx43266-283-B and c-Src close to the plasma membrane in the merge images. Please click here to view a larger version of this figure.
