As a demonstration of the value of this assay we have used it to investigate potential hypoxia in larvae with impaired function of the gene uninflatable (uif) in the tracheae. uif encodes a large transmembrane protein that is strongly expressed on the apical surface of the larval tracheal cells. Mutants of uif were previously noted to exhibit aberrant behavior that could indicate tissue hypoxia due to tracheal malfunction 9. We specifically suppressed uif expression in the larval trachea using the Gal4-UAS system. Two Gal4 lines were used: i) breathless (btl)-Gal4, which is expressed strongly in the tracheae from the beginning of their embryonic development and ii) cut(ue)-Gal4, which begins expression in a small posterior region of the main dorsal trunk tracheae at the end of embryogenesis and continues strong expression in this region throughout larval life 7. A UAS-uif RNAi line was obtained from the Vienna Drosophila Research Center (VDRC ID #1050).
As described in the protocol above, five replicate plates of newly hatched first instar larvae were set up for each of four crosses as follows.
Experiment 1
1) Control 1 – cut(ue)-Gal4 x Canton-S (+)
2) Experimental 1 – cut(ue)-Gal4 x UAS-uif RNAi
Experiment 2
3) Control 2 – btl-Gal4 x Canton-S (+)
4) Experimental 2 – btl-Gal4 x UAS-uif RNAi
The behavior of the cut(ue)-Gal4>UAS-uif RNAi larvae of Experiment 1 was comparable to that of the control cut(ue)-Gal4> + animals in terms of burrowing, tunneling and survival to pupation, (Figure 1 and Figure 2). In contrast, down-regulating uif expression throughout the entire tracheal system from early in development (Experiment 2) produced markedly different responses. The btl-Gal4>UAS-uif RNAi larvae displayed both behavioral manifestations of tissue hypoxia – reduced food burrowing and a total absence of substratum tunneling later in larval life (Figure 1 and Figure 2). The experimental larvae were also noticeably smaller, thinner and more sluggish than controls (Figure 3), and showed a high death rate during the course of the assay. As discussed above, reduced growth, sluggishness and organismal death are known symptoms of larval hypoxia. In addition, most of the experimental larvae failed to attempt pupation and remained as third instar larvae long after control larvae had pupated. Some larvae survived more than 15 days before eventually dying without pupating (Figure 1A). A few of the experimental larvae tried to pupate but in all cases abnormal pupae were formed that did not generate viable adults.
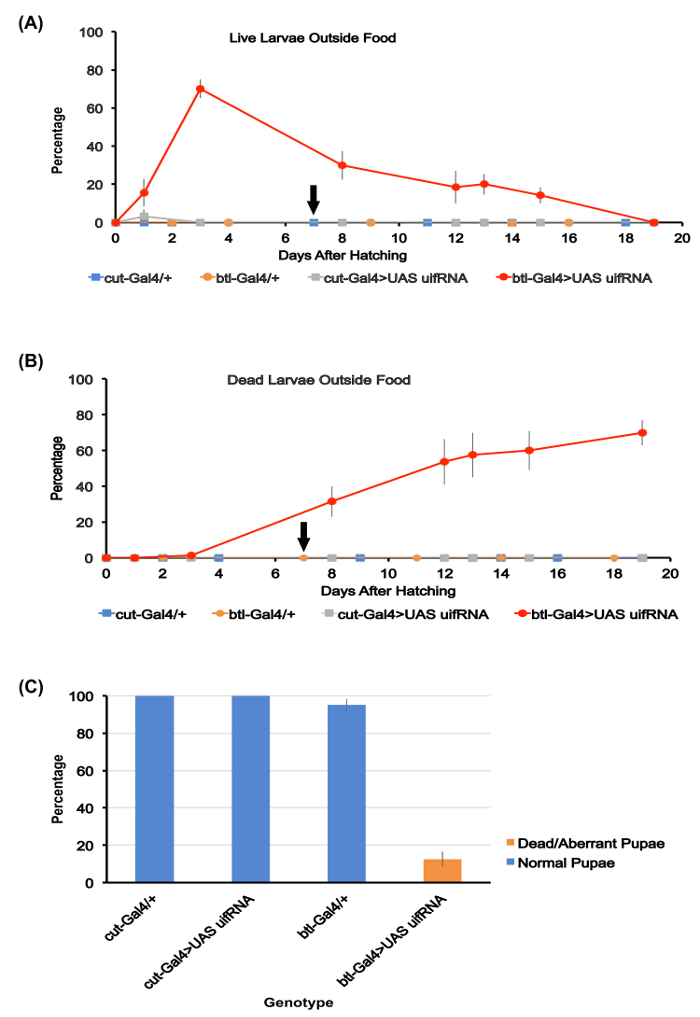
Figure 1. Quantitation of food burrowing and larval development.
(A) Percentage of live larvae outside the food mounds for the four genotypes studied here. Averages for the five assay plates used for each genotype are shown. By day 3 after hatching, ~70% of the btl-Gal4>UAS uif RNAi larvae were outside the food, whereas no larvae were detected outside the food or on the agar surface for the other three genotypes. The btl-Gal4>UAS uif RNAi larvae die slowly between days 4-20, with ~15% of them still alive 15 days after hatching. The black arrow along the x axis here and in (B) indicates the day by which all larvae of the three other genotypes had pupated.
(B) Percentage of dead larvae visible outside the food for the four genotypes. Averages for the five assay plates for each genotype are shown. Dead btl-Gal4>UAS uif RNAi larvae accumulate over time with most dying outside the food. The other genotypes all survive to pupation
(C) Percentage survival to pupation for the four genotypes studied. Averages for the five assay plates for each genotype are shown. No normal pupae of the btl-Gal4>UAS uif RNAi genotype were produced. Error bars throughout figure represent SEMs. Please click here to view a larger version of this figure.
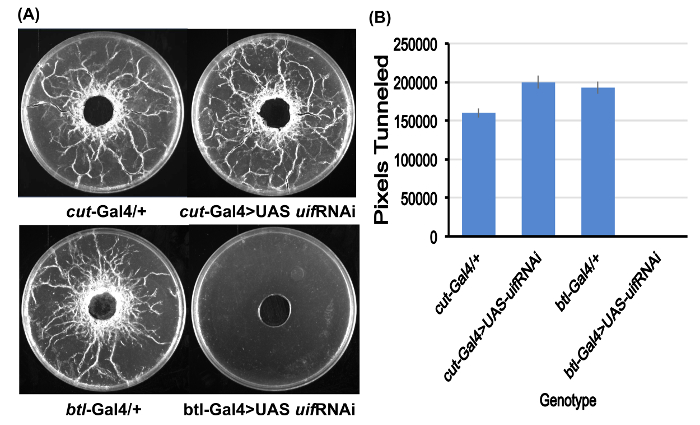
Figure 2. Quantitation of larval tunneling.
(A) Examples of the assay plates for all four genotypes studied after preparation for tunneling quantitation. Note complete absence of tunneling for the btl-Gal4>UAS uif RNAi larvae. Assay plates are 10 cm Petri plates.
(B) Tunneling quantitation derived from Image J for the four genotypes. Average pixel values for the five assay plates for each genotype. No tunneling was observed for the btl-Gal4>UAS uif RNAi larvae. Error bars = SEMs. Please click here to view a larger version of this figure.

Figure 3. Comparison of btl-Gal4>UAS uif RNAi and btl-Gal4> + larvae
Larvae of similar age (day 6 after hatching) for the btl-Gal4>UAS uif RNAi (A) and btl-Gal4> + (B) genotypes. Red arrows point to the dorsal trunk tracheae. Both larvae imaged at the same magnification. Scale bar = 0.5 mm. Please click here to view a larger version of this figure.
