Forced Salivation As a Method to Analyze Vector Competence of Mosquitoes
Summary
For efficient control of mosquito borne virus transmission, the knowledge of the vector potential of respective mosquitoes is of particular interest. We describe forced salivation as a method to analyze vector competence of Aedes albopictus and three different Culex taxa for the transmission of Zika virus.
Abstract
Vector competence is defined as the potential of a mosquito species to transmit a mosquito-borne virus (mobovirus) to a vertebrate host. Viable virus particles are transmitted during a blood meal via the saliva of an infected mosquito. Forced salivation assays allow determining the vector potential on the basis of single mosquitoes, avoiding the use of animal experiments. The method is suitable to analyze a large number of mosquitoes in one experiment within a short period of time. Forced salivation assays were used to analyze 856 individual mosquitoes trapped in Germany, including two different Culex pipiens pipiens biotypes, Culex torrentium as well as Aedes albopictus, which were experimentally infected with Zika virus (ZIKV) and incubated at 18 °C or 27 °C for two and three weeks. The results indicated the lack of vector competence of the different Culex taxa for ZIKV. In contrast, Aedes albopictus was susceptible to ZIKV, but only at 27 °C, with transmission rates similar to an Aedes aegypti laboratory colony tested in parallel.
Introduction
In 2015, Zika virus (ZIKV), a mosquito-borne virus (mobovirus) belonging to the family Flaviviridae, emerged in Columbia and Brazil and spread rapidly across the Americas and the Caribbean, causing an epidemic with notable numbers of associated clinical cases of microcephaly and Guillain–Barré syndrome1. Mosquitoes of the species Aedes aegypti and Aedes albopictus are considered the primary and secondary vectors of ZIKV2, but other species of the genus Aedes have also proven to have experimental vector competence3. In contrast to Aedes species, most of the Culex species tested so far are not able to transmit the virus4. Merely, the data for Culex quinquefasciatus provided inconclusive results3. At present, information is lacking on mosquito vector competence for ZIKV under moderate temperature conditions (<20 °C). However, this information is substantial for the risk assessments of possible spreads to regions with temperate climate such as Central Europe.
A competent vector for ZIKV is able to acquire, maintain, amplify and finally transmit the virus. Therefore, testing the mosquito bodies or parts of the bodies, including legs, midgut, head or salivary glands, for ZIKV is not sufficient to determine vector competence. It is mandatory to test for infectious virus particles within released saliva. To assess vector competence, both infection rate (IR, number of ZIKV-positive mosquito bodies per number of engorged mosquitoes) and transmission rate (TR, number of mosquitoes with ZIKV-positive saliva per number of ZIKV-positive mosquito bodies), have to be determined. IRs of mosquitoes can be easily analyzed by simply homogenizing mosquitoes followed by a reverse-transcriptase real-time polymerase chain reaction (RT-qPCR) targeting ZIKV. Moreover, the infection can be further characterized by determining the viral copy numbers per mosquito. In contrast, the analysis of transmission rates relies on the detection of viable virus particles in the saliva of individual mosquitoes. This can be achieved by feeding of infected mosquitoes on susceptible host animals, followed by the analysis of the viremia in the host5. However, this method relies on suitable model organisms, and is costly and restricted by animal-welfare regulations. To avoid the use of laboratory animals for analyzing transmission rates and to reduce costs, artificial re-feeding systems using blood droplets have been described6. However, testing on individual scale combined with a high number of individuals is difficult and time-consuming. The first description of a salivation assay on individual scale was published in 19667 and during recent years, forced salivation experiments have become the method of choice to evaluate the vector competence of mosquitoes3,8.
Based on our vector competence study of Central European mosquitoes published recently9, we describe forced salivation as reported by Anderson et al.8 with some modifications. This method allows high throughput standardized experiments under high biosafety level conditions (BSL-3) and includes the identification of active virus by cell culture based assays. The experiments described here include 856 mosquitoes. They were infected with ZIKV by artificial blood meal and subsequently analyzed for the presence of infectious virus particles in the saliva. The mosquitoes introduced into the experiments comprised two long-established laboratory populations of Ae. aegypti and Culex pipiens pipiens biotype molestus (Cx. p. molestus), as well as field caught Culex pipiens pipiens biotype pipiens (Cx. p. pipiens), Culex torrentium (Cx. torrentium) and two Ae. albopictus populations. With the exception of one of the two Ae. albopictus populations, which was collected in Italy, all mosquitoes were collected in Germany.
Protocol
1. Prepare Female Mosquitoes
- Collect mosquitoes from a big cage by using an aspirator (approximately 400 mosquitoes per big cage).
- Anesthetize the mosquitoes with carbon dioxide (CO2) for 7 s.
- Sort 20 females per plastic vial (Ø 50 mm × 100 mm).
Note: If the mosquitoes wake up, use another 3 s of CO2. - Starve the mosquitoes overnight.
2. Artificial Blood Meal
NOTE: All steps are performed under BSL-3 conditions.
- Prepare the infectious blood meal.
- Dilute the virus stock to the concentration of 1 x 108 plaque forming units (PFU)/mL.
Note: The stock concentration was determined by performing Tissue culture infection dose 50 method (TCID50) and calculated by the Spearman & Kärber algorithm10,11. - Mix expired human blood (blood preservation) from the blood bank (not suitable for human anymore, but useful for mosquitoes) with fructose (8% solution), filtrated bovine serum (FBS) and working solution of the virus stock (expired human blood:fructose:FBS:working solution = 5:3:1:1).
Note: We chose human blood, because this is the known natural mammal host of ZIKV.
- Dilute the virus stock to the concentration of 1 x 108 plaque forming units (PFU)/mL.
- Freeze 140 µL of blood meal mix for further analysis via TCID50.
- Perform artificial feeding by using different feeding methods, depending on the species:
- Aedes aegypti: Use a membrane feeding system for 30 min (1 mL per feeder).
- Culex spp.: Provide the infectious blood via a cotton stick overnight and do not warm up the blood (300 µL per vial).
- Aedes albopictus: Put two droplets (50 µL each) of the blood meal mix into the plastic vial and let the mosquitoes feed for 2 h. Do not warm up the blood.
Note: Mosquitoes will feed in the first 15 min. The prolonged incubation time is necessary due to safety reasons. The blood droplets are drained after 2 h, thus the contamination risk is reduced during the sorting of fed mosquitoes.
- Anesthetize the mosquitoes with CO2 for 7 s.
NOTE: If the mosquitoes wake up, use another 3 s of CO2. - Sort and count fully engorged mosquitoes into a new vial.
- Add a cotton pad, saturated with fructose (8% solution) between the vial and a plug.
- Keep the mosquitos at the designated temperature of 18 °C or 27 °C in the humidity of 80% for 14 or 21 days.
- Feed the mosquitos with fructose-saturated cotton pads. Replenish every 72 h with 1.5 mL of fructose (8% solution) per vial (up to 20 mosquitoes, depending on the feeding rate, see step 2.5).
3. Forced Salivation Assay on Day 14 or 21
NOTE: All steps are performed under BSL-3 conditions.
- Seed 2 x 104 Vero cells/per well in a 96 well plate with 200 µL of growth medium (Dulbecco´s modified eagle medium (DMEM) supplemented with 3% FBS, 100 U/mL penicillin, 100 mg/mL streptomycin, 5 µg/mL amphotericin B, 2 mM L-glutamine, 1% non-essential amino acids (NEAA), 1% sodium pyruvate).
- Prepare the salivation device:
- Place a squared glass plate (20 × 20 cm) on the bench in an angle of 30°.
Note: This is necessary due to the underpressure in the safety lab. If the plate is horizontal or vertical, liquid will leak. - Place double sided adhesive tape on top of the glass plate.
- Role the modeling clay until it reaches a diameter of 0.5 cm and attach it horizontally 1 cm away from the double-sided adhesive tape.
- Cut off the first 0.3 cm of the tip of a 10 µL filter tip.
NOTE: It is advised to prepare this before starting the experiment outside of the biosafety laboratory. - Fill the filter tips with 10 µL of phosphate buffered saline (PBS, pH 7.4).
- Place the filter tips on the modeling clay with the tip towards the sticky tape.
- Secure the filter tips with gentle pressure.
- Place a squared glass plate (20 × 20 cm) on the bench in an angle of 30°.
- Prepare the mosquitoes:
- Anesthetize the mosquitoes with CO2.
- Remove the legs and wings to immobilize the mosquitoes.
- Fix the living mosquito bodies on the sticky tape above a filter tip.
- Place the proboscis gently into the filter tip.
- Run the salivation assay for 30 min.
- Remove the filter tips and discard the clay and tape.
4. Processing of Saliva
NOTE: Steps 4.1–4.6.2 are performed under BSL-3 conditions.
- Expel the contents into the reaction tubes containing 10 µL of PBS.
- Mix gently and transfer the content into the wells of the prepared 96 well plate using one well per sample.
- Incubate the plate for 7 days at 37 °C with 5% CO2.
- Use the binocular to check the cells for the presence of cytopathic effect (CPE).
- If the well is positive for CPE, harvest the supernatant by pipetting 140 µL into a reaction tube for RNA extraction.
- Purify the RNA by using a RNA extraction kit.
- Mix 140 µL of the supernatant with 560 µL of AVL buffer (viral lysis buffer) and incubate for 10 min at room temperature.
- Add 560 µL of ethanol (96%) and vortex the sample.
NOTE: The samples are inactivated after this step and can be discharged out of the BSL-3 laboratory. - Pipet 630 µL of the sample onto a column and centrifuge with 6,000 x g for 1 min.
- Discard the filtrate, pipet the remaining 630 µL on the column and repeat the centrifugation.
- Wash the RNA, bound to the membrane of the column, by adding 500 µL of washing buffer 1 to the column and centrifuge at 6,000 x g for 1 min.
- Discard the collection tube and place the column into a new collection tube. Add 500 µL washing buffer 2 and centrifuge at 12,500 x g for 3 min.
- Discard the collection tube and place the column into a 1.5 mL reaction tube.
- Add 50 µL of elution buffer and incubate for 1 min at room temperature.
- Centrifuge at 6,000 x g for 1 min and then discard the column.
- Analyze the samples for ZIKV RNA via RT-qPCR.
5. Processing of Mosquito Bodies
NOTE: Steps 5.1–5.5 are performed under BSL-3 conditions.
- Remove the bodies of the mosquito and place each body into a reaction tube.
NOTE: It is possible to pause the experiment at this point and store the samples at -80 °C. - Add 500 µL of homogenization media (DMEM without any supplementals) into the reaction tube.
- Homogenize the body using hand motor mixer and use a new pistil for every sample.
- Add 200 µL of the homogenate into a 96-well-plate for purification.
NOTE: It is possible to pause the experiment at this point and store the samples at -80 °C. - Inactivate the plate by incubation for 60 min at 60 °C.
- Purify the RNA by automated nucleic acid purification.
NOTE: It is possible to pause the experiment at this point and store the samples at -80 °C.
6. Analysis
- Quantify the Viral RNA Copies using RT-qPCR.
NOTE: The RNA copies were averaged over all ZIKV-positive mosquito bodies and without including the ZIKV-negative mosquitoes. - Calculate the IR, defined as the number of ZIKV-positive mosquito bodies per number of fed females.
- Calculate the TR, defined as the number of mosquitoes with ZIKV-positive saliva per number of ZIKV-positive mosquito bodies. The transmission rate could not be calculated for species-temperature combinations with no ZIKV-positive bodies.
Representative Results
Aedes aegypti is known as a competent vector for ZIKV at least under tropical climatic conditions12. Therefore, we used Ae. aegypti as a positive control to establish the assay and to analyze IRs as well as TRs at an incubation temperature of 27 °C and a humidity of 80%. IR and TR were defined as previously described by Fortuna et al.13. Challenged Ae. aegypti showed IR of 50% and 72% and TR of 42% and 31% at 14 and 21 days post infection (dpi), respectively (Figure 1A and 1B). Subsequently, European mosquitoes were tested for ZIKV infection and transmission using tropical as well as temperate climatic conditions.
At 27 °C incubation temperature, all mosquito species showed ZIKV infection at 14 dpi as well as 21 dpi, except Cx. p. pipiens and Cx. torrentium individuals, which were positive at 14 dpi only. All Aedes species showed higher IRs (between 50% and 72%) as well as higher viral RNA concentration (106 up to 109 RNA copies/mosquito specimens) in comparison to the different Culex taxa, which revealed IRs ranging between 0% and 32% and viral loads of 103 to 104 RNA copies/mosquito specimens. All Aedes species displayed ZIKV-positive saliva (TR 13–42%), whereas none of the saliva samples from the Culex taxa contained ZIKV (Figure 1A and 1B). Interestingly, transmission rates at 21 dpi were similar in Ae. aegypti and Ae. albopictus from Germany (30%), but were substantially lower in Ae. albopictus from Italy (13%).
Incubation of challenged mosquitoes at a moderate temperature of 18 °C revealed generally lower virus loads in the Aedes species (104–106 RNA copies/mosquito) as well as in the different Culex taxa (102–104 RNA copies/mosquito). However, at 18 °C incubation temperature, transmission of ZIKV was not observed for any of the mosquito species investigated.
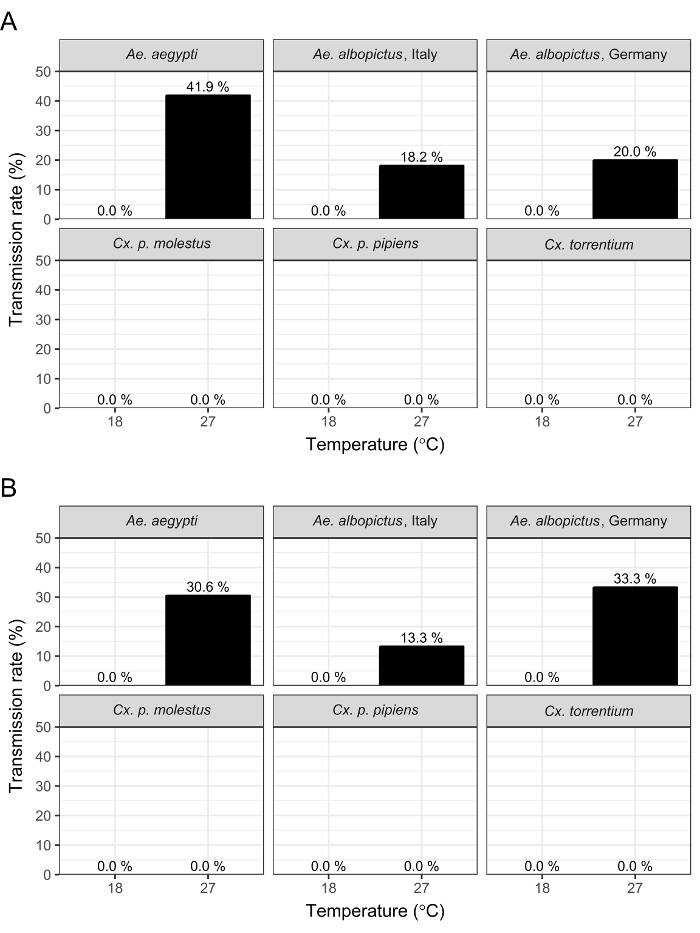
Figure 1: Transmission rates of mosquitoes experimentally infected with ZIKV. Results of vector competence studies with ZIKV in different mosquito species. Six different mosquito taxa were infected with ZIKV and incubated at 27 °C or 18 °C for 14 (A) or 21 (B) days. TR is defined as the number of mosquitoes with ZIKV-positive saliva per number of ZIKV-positive mosquito bodies. The total number of mosquitoes investigated was 856, with a minimum of 30 individuals for each time point/temperature/mosquito taxa. Please click here to view a larger version of this figure.
Discussion
It has previously been shown that results obtained with forced salivation are in line with classical re-feeding experiments6. However, both in re-feeding experiments and in our presented method of forced salivation, it is impossible to directly proof salivation activity or monitor the release of saliva. To prove salivation activity, samples could be tested for other components present in the saliva (e.g., proteins, carbohydrates, etc.). Furthermore, additional qPCRs would allow detection of viral RNA copies. This, however, requires splitting of saliva samples for various analyses, which may limit sensitivity in the cell culture assay in case of very low particle numbers. In turn, this might lead to an underestimation of the determined transmission rates.
For reproducible results, it is necessary to ensure that all mosquitoes stay alive until the end of the experiment. This is controlled by visually monitoring the body movement activity of the mosquitoes. Furthermore, the principle use of the salivation assay for a given mosquito species has to be tested by feeding control viruses, which are known to be transmitted by this specific mosquito species. Vice versa, every virus introduced into an experiment has to be tested in a susceptible mosquito.
Forced salivation assays have many advantages. High numbers of mosquitoes can be tested simultaneously on a single specimen base under controlled standardized conditions without conflicting with animal welfare regulations9.
The method is used by several laboratories. However, exact setups may differ, which can lead to differences in results between laboratories. One critical component is the capillary to collect the saliva, which can be made of glass14 or plastic15. To comply with the high safety regulations of a biosafety level 3 insectary, we used plastic filter tips instead of glass capillaries, thus reducing the risk of injuries. Furthermore, filter tips have the advantage that the collected liquid can easily be transferred using a pipette.
The setup of the forced salivation assay presented here is based on two investigators working together and sharing duties. Demobilizing of the mosquito is performed by one person, while the other simultaneously prepares the forced salivation setup. This significantly reduces handling time and minimizes the risk of sample switching as changing between the demobilization-workplace and the salivation-plate is not necessary. Moreover, it allows placing the mosquitoes on the forced salivation device immediately after demobilization. This rather short mosquito handling time is essential for successful forced salivation. Lastly, the mosquitoes can be directly monitored for motility throughout the whole experiment.
The salivation assay presented here is used to gain insights into the vector potential of mosquito species. However, to answer some other specific questions (e.g., the numbers of mosquito bites that are needed to infect vertebrates), animal experiments might still be required.
Disclosures
The authors have nothing to disclose.
Acknowledgements
We thank Ella Weinert, Michelle Helms and Marlis Badusche for excellent technical assistance, Jessica Börstler for analyses of Culex mosquitoes, Norbert Becker and Björn Pluskota for providing Aedes albopictus eggs and Olli Vapalahti for providing the virus stock. ML was supported by the Leibniz Association; grant number SAW-2014-SGN-3.
Materials
| Inject+matic Sleeper | Inject+matic | ||
| Bugdorm-1 Insect Rearing Cage | BugDorm Store | ||
| Drosophila cultivation tubes | carl roth | PK11.2 | plastic vial for mosquitoes |
| Plug for drosophila cultivation tubes | carl roth | PK15.1 | |
| Constant climate chamber | Binder | KBF-240 | |
| Blood | banked human blood, expired, not usable for medical application | ||
| Dumont-Pincette Electronic SS | Dumont | B40c | |
| Vero cells | BNITM | ||
| Flash light aspirator | Bioquip | 2809 | |
| double-sided adhesive tape | no specific provider | ||
| Gammex Powder-free gloves with AMT | Ansell | for safety reasons necessary during work with pricks in the BSL3 laboratory | |
| RealStar Zika Virus RT-PCR-Kit | altona diagnostics | 591013 | |
| MagMax Pathogen RNA/DNA Kit | Thermo fisher scientific | 4462359 | RNA isolation of mosquito bodies |
| Qiamp viral RNA Mini Kit | Qiagen | 52906 | RNA-Isolation of supernatant |
| filter tips (20 µL) | Sarstedt | 701,116,210 | Cut the first 3 mm of the tip |
| hand motor mixer | carl roth | sold out | |
| micro pestles for hand motor mixer | carl roth | CXH8.1 | |
| DMEM | P0403550 | ||
| L Glutamine Penicilin/Streptomycin | PAN | P0819100 | |
| Amphotericin B | PAN | P06-0110 | |
| Fructose | carl roth | 4981.5 | |
| Phosphate buffered saline | PAN | P04-36500 | |
| FBS | PAN | ||
| Sodium Pyruvat | PAN | P0443100 | |
| MEM NAA | PAN | P0832100 | |
| Hemotek Feeder | Hemotek | PS6 | |
| Light Cycler | Roche | 480 II | |
| Light Cycler Software | Roche | 480 SW 1.5.1 | |
| Modeling clay | no specific provider | ||
| King Fisher Flex Purification System | ThermoFisher scientific | ||
| Aedes albopictus | eggs were collected in Freiburg, Germany | ||
| Binocular | MOTIC | SMZ-168 | |
| Binocular light | MOTIC | MLC-150C | |
| CO2-Incubator | Thermo scientific | MIDI 40 |
References
- Musso, D., Gubler, D. J. Zika Virus. Clinical Microbiology Reviews. (3), 487-524 (2016).
- Epelboin, Y., Talaga, S., Epelboin, L., Dusfour, I. Zika virus: An updated review of competent or naturally infected mosquitoes. PLOS Neglected Tropical Diseases. 11 (11), e0005933 (2017).
- Aliota, M. T., Peinado, S. A., Osorio, J. E., Bartholomay, L. C. Culex pipiens and Aedes triseriatus Mosquito Susceptibility to Zika Virus. Emerging Infectious Diseases. (10), 1857-1859 (2016).
- Turell, M. J., Linthicum, K. J., Patrican, L. A., Davies, F. G., Kairo, A., Bailey, C. L. Vector competence of selected African mosquito (Diptera: Culicidae) species for Rift Valley fever virus. Journal of Medical Entomology. 45 (1), 102-108 (2008).
- Cornel, A. J., Jupp, P. G. Comparison of three methods for determining transmission rates in vector competence studies with Culex univittatus and West Nile and Sindbis viruses. Journal of the American Mosquito Control Association. 5 (1), 70-72 (1989).
- Hurlbut, H. S. Mosquito salivation and virus transmission. The American Journal of Tropical Medicine and Hygiene. 15 (6), 989-993 (1966).
- Anderson, S. L., Richards, S. L., Smartt, C. T. A simple method for determining arbovirus transmission in mosquitoes. Journal of the American Mosquito Control Association. 26 (1), 108-111 (2010).
- Heitmann, A., Jansen, S., Lühken, R., Leggewie, M., Badusche, M., Pluskota, B., et al. Experimental transmission of Zika virus by mosquitoes from central Europe. Eurovsurveillance. 22 (2), (2017).
- Hierholzer, J. C., Killington, R. A. . Virus Isolation and Quantitation – Virology methods manual. , (1996).
- Horzinek, R. W., Marian, C. Kompendium der allgemeinen Virologie. Zentralblatt für Veterinärmedizin R A. 32 (1-10), 480 (2010).
- Richard, V., Paoaafaite, T., Cao-Lormeau, V. -. M. Vector Competence of French Polynesian Aedes aegypti and Aedes polynesiensis for Zika Virus. PLOS Neglected Tropical Diseases. 10 (9), (2016).
- Fortuna, C., Remoli, M. E., Di Luca, M., Severini, F., Toma, L., Benedetti, E., et al. Experimental studies on comparison of the vector competence of four Italian Culex pipiens populations for West Nile virus. Parasites & Vectors. 8, 463 (2015).
- Weger-Lucarelli, J., Rückert, C., Chotiwan, N., Nguyen, C., Garcia Luna, S. M., Turell, M. J., et al. Vector Competence of American Mosquitoes for Three Strains of Zika Virus. PLOS Neglected Tropical Diseases. 10 (10), (2016).
- Dubrulle, M., Mousson, L., Moutailler, S., Vazeille, M., Failloux, A. -. B. Chikungunya virus and Aedes mosquitoes: saliva is infectious as soon as two days after oral infection. PLOS ONE. 4 (6), e5895 (2009).
