In this protocol, Aβ1-42 was incubated at 37 °C for 7 days before use, because incubation of Aβ1-42 was needed for producing toxic forms27,28,30,35. After this incubation, aggregated forms of Aβ were observed (Figure 1A). It has been reported that similar incubation of Aβ1-42 produced the fibril form of Aβ36. After treatment with this aggregated Aβ1-42, immunostaining with an antibody for the toxic oligomer of Aβ35,37 was performed, and positive staining was detected on cultured neurons (Figure 1B). Considering the above, this incubation protocol produces the toxic forms of Aβ.
Several days were required for the induction of axonal degeneration after Aβ exposure. The events prior to axonal degeneration remain unclear. Therefore, this protocol has been developed to further understand the mechanisms involved. Using this protocol, the early phenomena induced by Aβ treatment were analyzed. Cortical neurons were cultured for 4 days. The longest neurites in the cultured neurons were identified as axons; these were confirmed by positive immunostaining for the axonal marker, tau-1, and negative immunostaining for the dendritic marker, MAP2 (Figure 2). After 1 h of vehicle treatment, growth cones had spread lamellipodia and processed several filopodia. These were identified as healthy growth cones. Conversely, 1 h of Aβ1-42 treatment led to shrunken growth cones, which developed no lamellipodia or filopodia. These were identified as collapsed growth cones. Collapse scores were calculated as described in step 1.3.7. When shapes of growth cones were unclear, they were eliminated from the analysis. Aβ1-42 treatment led to a significant increase in collapse score, corresponding to increased axonal growth collapse, when compared to the collapse score of vehicle-treated growth cones27.
Axonal growth cones were observed before and after treatment with Aβ1-42 (Figure 3). Cells were maintained in the inverted microscope with a humidified atmosphere of 10% CO2 at 37 °C. Images were captured every 5 min. As shown in Figure 3, growth cones collapsed between 21 and 26 min after Aβ1-42 treatment. Growth cones were excluded from live cell imaging if they did not retain their healthy shape for 1 h prior to any treatment.
To visualize the early effects of Aβ1-42-treatment, endocytosis was used as the focus of this analysis, because endocytosis inhibitors can block Aβ1-42-induced growth-cone collapse27. Endocytosis was visualized with a fluorescence membrane probe (i.e., a fluorescent dye that binds to plasma membranes and is spontaneously endocytosed). A previous study showed that growth cones do not collapse at 20 min after Aβ1-42-treatment27; therefore, healthy growth cones were selected by DIC imaging in vehicle- or Aβ1-42-treated cells after 20 min. Following Aβ1-42-treatment, numerous fluorescent membrane probe-positive puncta were observed in the growth cone (Figure 4). The density of fluorescence membrane probe-positive puncta in growth cones was significantly increased27. This suggests that Aβ1-42-induced growth cone endocytosis occurs prior to collapse.
To confirm the role of endocytosis, a DNA plasmid encoding EGFP-AP180 C-terminus was transfected into cultured cortical neurons. Cells expressing the AP180 C-terminus selectively inhibited clathrin-mediated endocytosis38,39. If EGFP expression was observed at the cell body in the neuron, the AP180 C-terminus was considered to be expressed at the axonal growth cone of the neuron. Transfection of AP180 C-terminus blocked Aβ1-42-induced growth cone collapse (Figure 5)27.
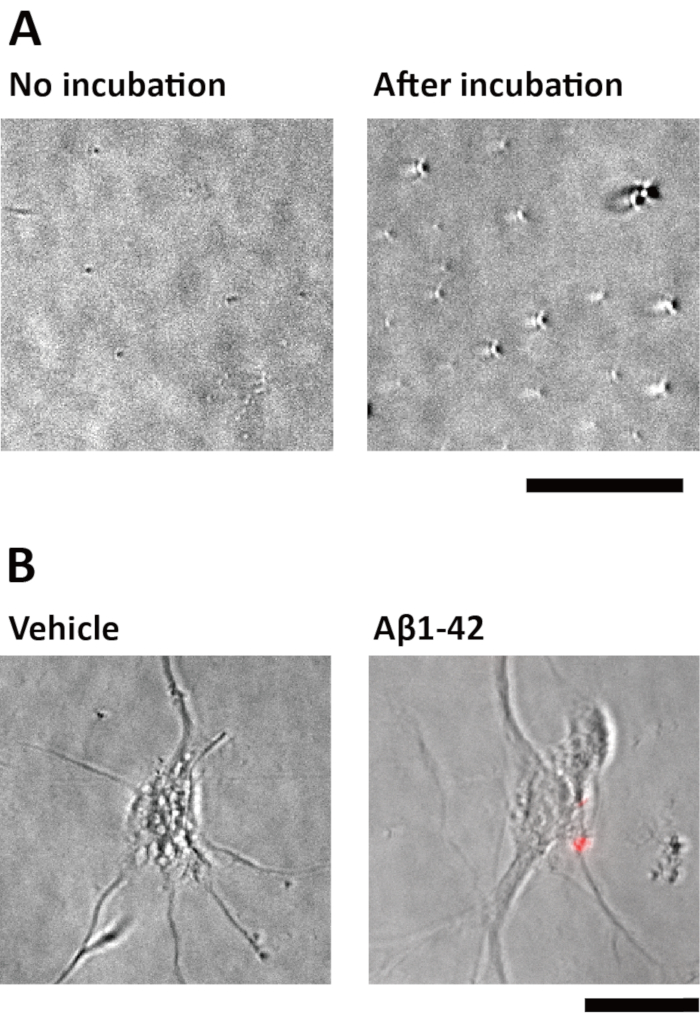
Figure 1: Incubation of Aβ1-42 aggregates Aβ. (A) Aβ1-42 was dissolved in distilled water at a concentration of 0.5 mM and incubated at 37 °C for 7 days (after incubation), or stored at -30 °C without incubation (no incubation). Each Aβ solution was diluted to 0.1 mM; then, 10 μL of each diluted solution was dropped on glass slides and covered with coverslips. Bright-field images with oblique illumination were captured by using inverted microscope B. Scale bar = 20 μm. (B) Aggregated Aβ1-42 or vehicle treatment on cultured neurons for 4 h. Following treatment, the neurons were fixed and immunostained for toxic Aβ oligomers. Fluorescence images (red) and bright-field images with oblique illumination (gray) are shown. Scale bar = 20 μm. Please click here to view a larger version of this figure.

Figure 2: Aβ1-42-induced axonal growth cone collapse. After Aβ1-42- or vehicle-treatment, neurons were fixed and immunostained for tau-1 (red) and microtubule associated protein 2 (MAP2, green). Fluorescence and differential interference contrast (DIC) images are shown. Magnified views of the regions of interest (ROI, rectangles) are shown below their corresponding images. White scale bars = 50 μm; black scale bars = 10 μm. This figure has been modified from Kuboyama et al, 201527. Please click here to view a larger version of this figure.
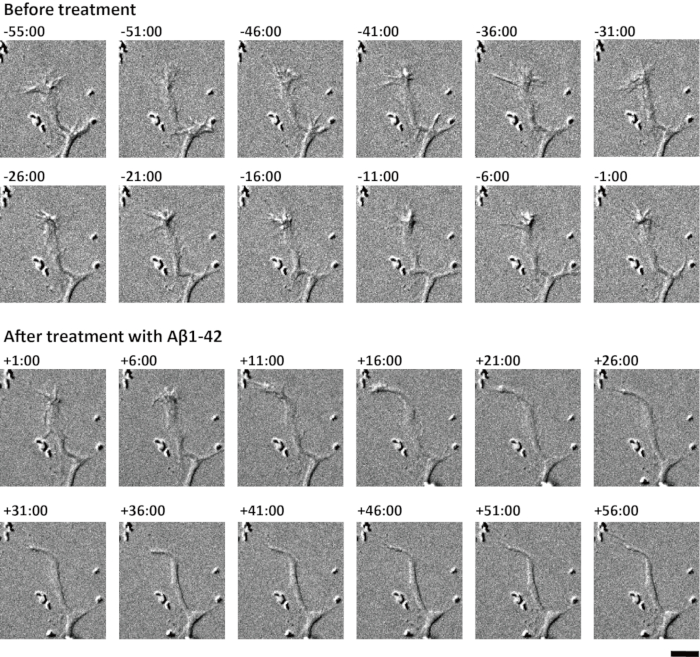
Figure 3: Live cell imaging before and after Aβ1-42 treatment. After 4 days of culture, cells were transferred to an inverted microscope and DIC images were captured every 5 min. Time-lapse images are shown. The digits represent minutes:seconds after the application of aggregated Aβ1-42 (final concentration, 0.5 μM). Scale bar = 10 μm. Please click here to view a larger version of this figure.
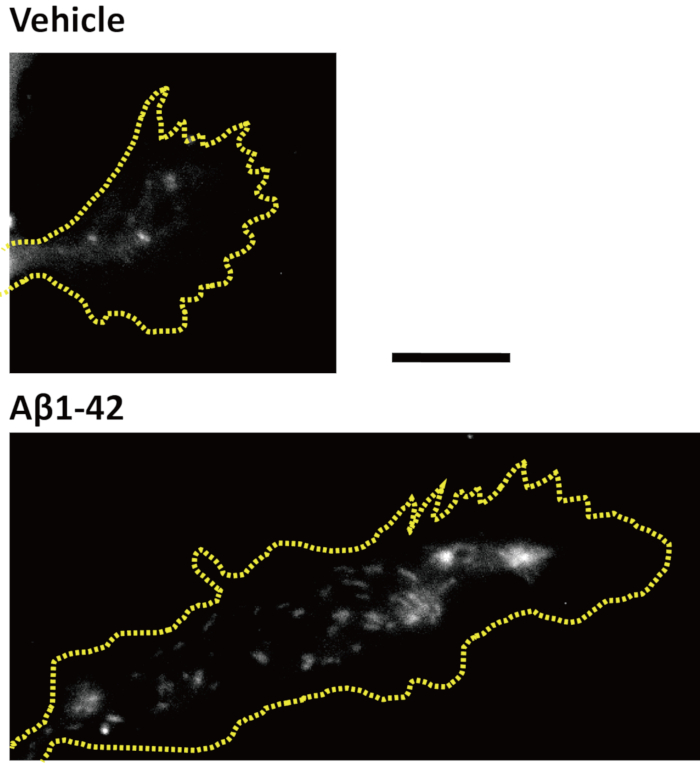
Figure 4: Twenty minutes of Aβ1-42 treatment induced endocytosis. Cortical neurons were cultured for 4 days and treated with a fluorescence membrane probe. Then, neurons were treated for 20 min with Aβ1-42 or vehicle. Fluorescence images of the growth cones are shown. The yellow dotted lines represent the outlines of the growth cones. Scale bar = 10 μm. Please click here to view a larger version of this figure.
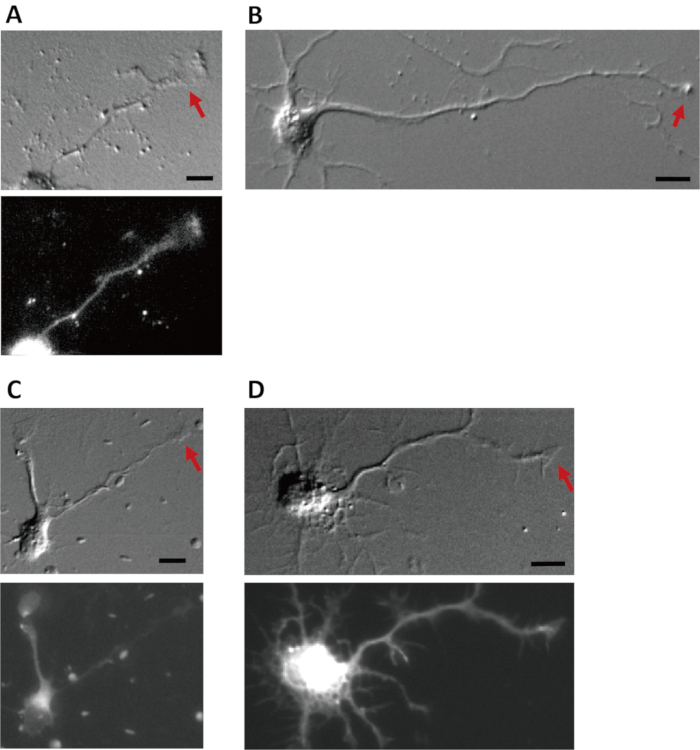
Figure 5: Expression of AP180-C terminus blocks Aβ1-42-induced collapse. Four days after transfection of EGFP (A, B) or EGFP-AP180 C-terminus (C, D); Aβ1-42 (B, D) or vehicle (A, C) was added to cortical neurons for 1 h. DIC (upper panels) and fluorescence (bottom panels) images are shown. Arrows indicate growth cones. Scale bars = 10 μm. Please click here to view a larger version of this figure.
