A pyoverdine mixture biosynthesized by Pseudomonas fluorescens was used as a standard to interpret and quantify absorbance (at 420 nm) of samples in terms of pyoverdine equivalents in µM. Figure 1 shows the relationship between absorbance (420 nm) and starting concentration of pyoverdine (Log10 molarity in µM). EDTA did not provide an adequate standard because samples exhibited greater absorbance measurements than were attainable with pyoverdine, and the R2 was lower (Figure 2). While initial work using the CAS-Fe assay as a method of siderophore detection measured absorbance at 630 nm, in a related study using a very similar method (CAS-Fe-Agar was mixed 1:1 with modified M9 to generate a 200 µL column in the microplate), it was observed that the peak absorbance was at 665 nm, but that 420 nm was more reproducible in terms of changes in absorbance induced by samples (Figure 3).
Siderophore production was observed in enrichment cultures of all tissue types after 72 h of Fe-deficit enrichment and siderophore activity appeared to stabilize after 48 h of incubation (Supplementary Figure 1). Thus, siderophore activity of the 72 h enrichment was assessed at 48 h incubation to determine the influence of genotype and sample type on siderophore isolation (Figure 4). Siderophore activity in bulk soil samples was relatively low and did not exhibit differences between the wheat genotype from which the bulk soil was sampled (Figure 4A). Enrichments of loosely bound soil isolated from the PI561725 genotype exhibited greater siderophore production compared with loosely bound soil from Madsen and PI561727, but not Lewjain (Figure 4B). Siderophore production in enrichments from tightly bound soil was not heavily influenced by genotype (Figure 4C).
Enrichment cultures of grain tissue yielded relatively low siderophore production regardless of genotype (Figure 4D). Enrichments of Lewjain shoot tissue had significantly lower siderophore production than the other genotypes, and PI561725 shoot tissue cultures resulted in more variable siderophore production (Figure 4E). Siderophore activity was more than 200% greater in in root tissue enrichment cultures of PI561725 compared with all other genotypes (Figure 4F).
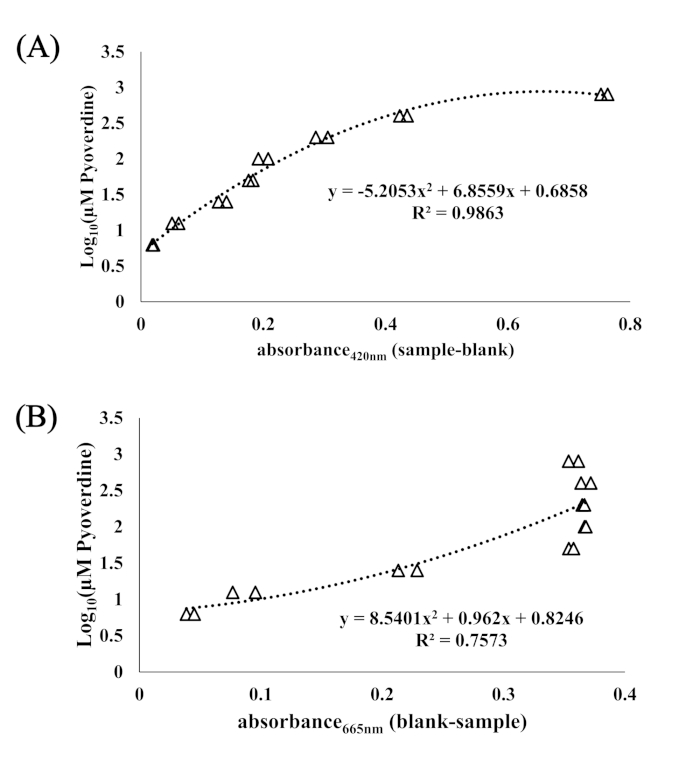
Figure 1. Absorbance at 420 nm and 655 nm regressed against the log10 concentration of pyoverdine. (A) Absorbance at 420 nm regressed against the log10 concentration of pyoverdine in µM. A polynomial curve was fit to obtain an explanatory equation for interpreting absorbance in terms of pyoverdine equivalents. (B) Absorbance at 665 nm regressed against the Log10 concentration of pyoverdine in µM. R2 is the square of the Pearson correlation coefficient, and the equation explains the fitted curve. Points are duplicates of absorbance measurements at 800, 400, 200, 100, 50, 25, 12.5, and 6.25 µM pyoverdine after 6 h incubation at 28 °C. Please click here to view a larger version of this figure.
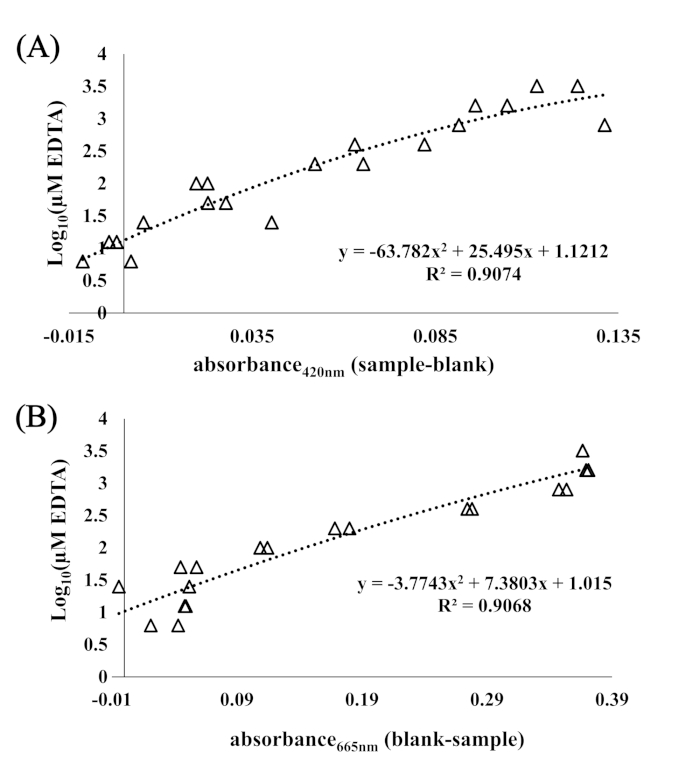
Figure 2. Absorbance at 420 nm and 655 nm regressed against the log10 concentration of EDTA. (A) Absorbance at 420 nm regressed against the log10 concentration of EDTA in µM. A polynomial curve was fit to obtain an explanatory equation for interpreting absorbance in terms of pyoverdine equivalents. (B) Absorbance at 665 nm regressed against the Log10 concentration of EDTA in µM. R2 is the square of the Pearson correlation coefficient, and the equation explains the fitted curve. Points are duplicates of absorbance measurements at 3200, 1600, 800, 400, 200, 100, 50, 25, 12.5, and 6.25 µM EDTA after 6 h incubation at 28 °C, and error bars . Please click here to view a larger version of this figure.
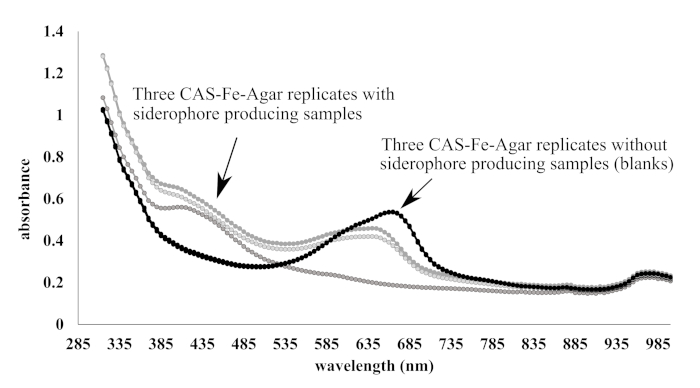
Figure 3. Absorbance scans from 315−1000 nm of microplate wells containing 200 µL columns of 1:1 CAS-Fe-Agar and modified M9 or M9 medium with siderophore producing samples. The plate was incubated at 28 °C for 72 hours before measuring absorbance in a microplate reader. Absorbance scans show the three blanks containing no sample (black lines) yielded tightly clustered curves with a peak at 665 nm. Absorbance scans show the three blanks containing siderophore producing samples (gray lines) yielded curves with more variability, but with more consistent absorbance at 420 nm compared with 665 nm. Please click here to view a larger version of this figure.
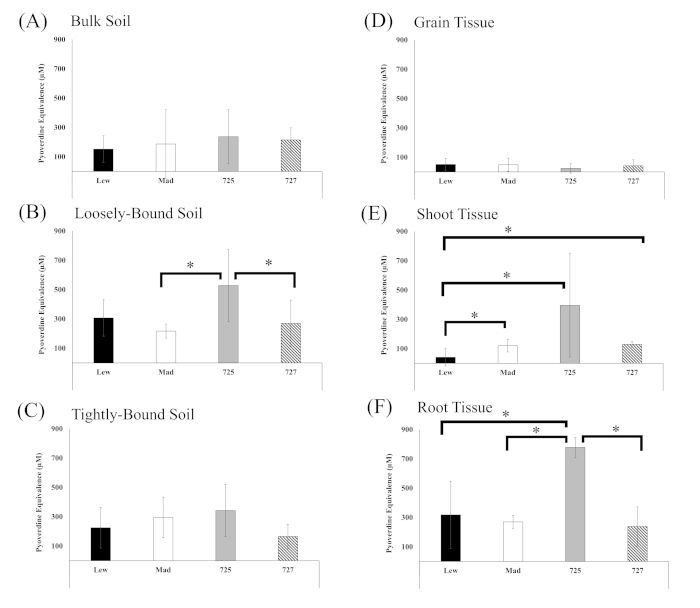
Figure 4. Pyoverdine equivalents of siderophore enrichment cultures. Pyoverdine equivalents of siderophore enrichment cultures associated with (A) bulk (B) loosely bound, and (C) tightly bound soil, and in tissue homogenates of wheat (D) grain (E) shoots, and (F) roots. Siderophore enrichment cultures were incubated for 72 h before transferring subsamples to a microplate and incubating at 28 °C. Siderophore production was assessed after 48 h of incubation with Chrome azurol S. Genotypes/Lines are Lew = Lewjain, Mad = Madsen, 725 = PI561725, and 727 = PI561727. Asterisks represent significance at alpha = 0.008 (after Bonferroni correction). Bars are standard deviation. Please click here to view a larger version of this figure.
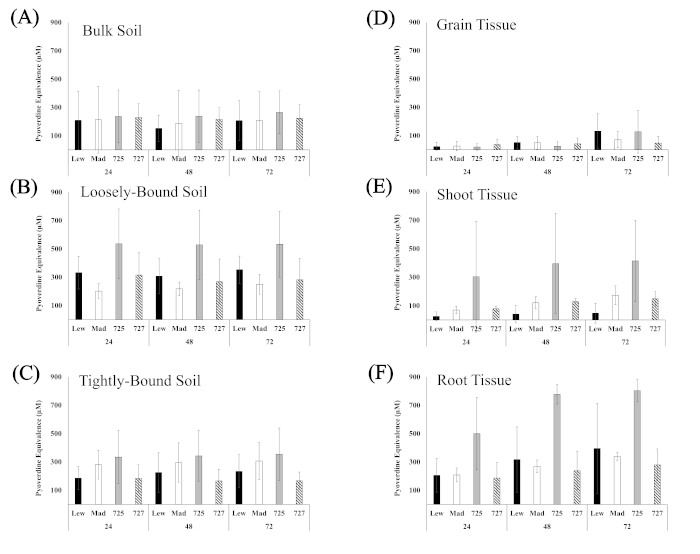
Supplementary Figure 1. Pyoverdine equivalents over time. Pyoverdine equivalents of siderophore enrichment cultures assessed after 24, 48, and 72 h of incubation with Chrome azurol S. Sderophore enrichment cultures associated with (A) bulk (B) loosely bound, and (C) tightly bound soil, and in tissue homogenates of wheat (D) grain (E) shoots, and (F) roots. Siderophore enrichment cultures were incubated for 72 h before transferring subsamples to a microplate and incubating at 28 °C. and subsampled to assess siderophore production at each timepoint. Genotypes/Lines are Lew = Lewjain, Mad = Madsen, 725 = PI561725, and 727 = PI561727. Siderophore production was assessed after, 24, 48, and 72 h of incubation. Bars are standard deviation. Please click here to view a larger version of this figure.
