Swimming Induced Paralysis to Assess Dopamine Signaling in Caenorhabditis elegans
Summary
Swimming induced paralysis (SWIP) is a well-established behavioral assay used to study the underlying mechanisms of dopamine signaling in Caenorhabditis elegans (C. elegans). However, a detailed method to perform the assay is lacking. Here, we describe a step-by-step protocol for SWIP.
Abstract
The swimming assay described in this protocol is a valid tool to identify proteins regulating the dopaminergic synapses. Similar to mammals, dopamine (DA) controls several functions in C. elegans including learning and motor activity. Conditions that stimulate DA release (e.g., amphetamine (AMPH) treatments) or that prevent DA clearance (e.g., animals lacking the DA transporter (dat-1) which are incapable of reaccumulating DA into the neurons) generate an excess of extracellular DA ultimately resulting in inhibited locomotion. This behavior is particularly evident when animals swim in water. In fact, while wild-type animals continue to swim for an extended period, dat-1 null mutants and wild-type treated with AMPH or inhibitors of the DA transporter sink to the bottom of the well and do not move. This behavior is termed “Swimming Induced Paralysis” (SWIP). Although the SWIP assay is well established, a detailed description of the method is lacking. Here, we describe a step-by-step guide to perform SWIP. To perform the assay, late larval stage-4 animals are placed in a glass spot plate containing control sucrose solution with or without AMPH. Animals are scored for their swimming behavior either manually by visualization under a stereoscope or automatically by recording with a camera mounted on the stereoscope. Videos are then analyzed using a tracking software, which yields a visual representation of thrashing frequency and paralysis in the form of heat maps. Both the manual and automated systems guarantee an easily quantifiable readout of the animals’ swimming ability and thus facilitate screening for animals bearing mutations within the dopaminergic system or for auxiliary genes. In addition, SWIP can be used to elucidate the mechanism of action of drugs of abuse such as AMPH.
Introduction
Animals perform a variety of innate and complex behaviors that are mediated by different neurotransmitters coordinated by intricate signaling processes. The neurotransmitter dopamine (DA) mediates highly conserved behaviors across species, including learning, motor function and reward processing.
The soil nematode C. elegans, with a relatively simple and well mapped nervous system consisting of only 302 neurons, shows markedly complex behaviors, including many that are regulated by DA such as mating, learning, foraging, locomotion and egg laying1. Among other features, short life cycle, ease of handling and the conservation of signaling molecules, highlight the advantages of using C. elegans as a model for studying the neural basis of conserved behaviors.
The hermaphrodite C. elegans contains eight dopaminergic neurons; In addition to these, the male contains six extra pairs for mating purposes. As in mammals, these neurons synthesize DA and express the DA transporter (DAT-1), a membrane protein found exclusively in dopaminergic neurons, which transports DA released in the synaptic cleft back into the dopaminergic neurons. Moreover, most of the proteins involved in each step of synthesis, packaging and release of DA are highly conserved between worms and humans and, like in mammals, DA modulates feeding behaviors and locomotion in C. elegans2.
C. elegans crawls on solid surfaces and swims with a characteristic thrashing behavior in water. Interestingly, mutants lacking expression of DAT-1 (dat-1) crawl normally on solid surface but fail to sustain swimming when immersed in water. This behavior was termed swimming induced paralysis, or SWIP. Previous experiments demonstrated that SWIP, in part, is caused by an excess of DA in the synaptic cleft that ultimately overstimulates the D2-like postsynaptic receptors (DOP-3). Although originally identified in dat-1 knockout animals3, SWIP is also observed in wild-type animals treated with drugs that block the activity of DAT (e.g., imipramine4) and/or induce DA release (e.g., amphetamine5). On the other hand, pharmacological or genetic manipulations averting synthesis and release of DA and blocking DOP-3 receptor function prevent SWIP6. Taken together, these already published data have established SWIP as a reliable tool to study the behavioral effects caused by mutated proteins within dopaminergic synapses3,4,7 and to be employed for forward genetic screens for the identification of novel regulatory pathways involved in DA signaling7,8,9,10,11,12. Additionally, by providing an easily quantifiable readout of drug-induced behavior in living animals, SWIP enables the elucidation of mechanisms of action of drugs like amphetamine (AMPH) and azaperone at the dopaminergic synapses5,6,13,14,15.
Protocols for performing the SWIP assays have been described before16. Here, we describe in detail the methodology and setup to perform the assay with the goal of providing a visual guide for the C. elegans community to effectively perform SWIP.
Protocol
1. Preparation of Solutions and Media
- Prepare M9 buffer by dissolving KH2PO4 3.0 g (22.05 mM), Na2HPO4 6.0 g (42.2 mM), and NaCl 5.0 g (85.5 mM) in 1 L of autoclaved deionized water. Add 1.0 mL of 1 M MgSO4 (12 g in a final volume of 100 mL autoclaved deionized water) after autoclaving. Mix 100 mL of the resulting 10x M9 with 900 mL of autoclaved deionized water to make a 1x solution.
- To make egg buffer, dissolve 6.896 g of NaCl (118 mM), 3.578 g of KCl (48 mM), 0.294 g of CaCl2-2H2O (2 mM), 0.406 g of MgCl2-6H2O (2 mM) and 5.958 g (25 mM) of HEPES in 1 L of autoclaved deionized water. Adjust pH to 7.3 using NaOH.
- Prepare fresh sodium hypochlorite/NaOH solution by adding 1 mL of 5-6% sodium hypochlorite (bleach) and 180 µL of 10 N NaOH to 3.8 mL of deionized water.
- Weigh 60 g sucrose and dissolve in autoclaved deionized water to a final volume of 100 mL to make 60% sucrose solution.
- Dissolve 0.684 g of sucrose in 10 mL of autoclaved deionized water to make 200 mM sucrose. Check and adjust to the same an osmolarity using osmometer. Make 1 mL aliquots in 1.5 mL microcentrifuge tubes and freeze at -20 °C.
- Weigh 0.184 g of AMPH (molecular weight 184.75 g/mol) and dissolve in 10 mL of deionized water to make a 100 mM stock solution. Mix 2 µL of the stock solution in 400 µL of water to make 0.5 mM working solution.
- Prepare Nutrient growth media (NGM) plates
- Mix 3 g of NaCl (52.65 mM), 20 g of peptone, 25 g of bacto-agar and 975 mL of deionized water in a 2 L Erlenmeyer flask. Include a magnetic stir bar and autoclave (121 °C, 15 PSI) for 1 hour using liquid cycle.
- Cool to and maintain the temperature at about 50 °C by placing the flask on a heater while stirring. Add 0.5 mL of cholesterol (5 mg/mL in ethanol), 1 mL of 1 M MgSO4, 1 mL of 1 M CaCl2 and 25 mL of 1 M potassium phosphate buffer, pH 7.4 (108.3 g of KH2PO4, 35.6 g of K2HPO4, deionized water to 1 L).
- Pipette 25 mL each into 100 mm x 15 mm Petri plates and allow the media to solidify. Store the plates upside down at 4 °C in a box for up to 4 weeks.
- Preparation of Lysogeny broth (LB) broth
- Dissolve 5 g of LB powder mix in 200 mL of deionized water in an Erlenmeyer flask. Autoclave for 30 minutes utilizing the liquid sterilization cycle. Allow the broth to cool down. Store at room temperature for 1-2 weeks.
- Preparation of NA22 bacterial plates
- Use a sterile pipette tip or a sterile bacterial loop to streak an LB plate with a small volume of NA22 E. coli bacteria from glycerol stock and incubate the plate upside down in a 37 °C incubator overnight to grow isolated colonies. Pick and introduce a single colony into 200 mL of LB broth prepared in step 1.8 and let grow overnight at 37 °C on a shaking platform.
- To seed the plates, dispense 200 µL of bacterial culture onto the NGM plates prepared earlier in step 1.7 and spread with a sterile glass hockey stick. Let the plates dry overnight or longer under a hood and store upside down in an airtight box at 4 °C.
- To make the eyelash/platinum tool to pick worms, glue a thick eyelash or a platinum filament into a glass Pasteur pipette using super glue. Cut the tip of the eyelash at an angle using a razor blade. Alternatively, a Bunsen burner can be used to melt the tip of the glass pipette around the platinum filament.
2. C. elegans Husbandry
NOTE: Culture wild-type N2C. elegans strain on Escherichia coli NA22 plates. The detailed culture methods are described below.
- Preparation of worm culture
- To make a starter culture of worms, cut a small piece of agar from a plate containing well-fed animals and transfer it onto a NA22 E. coli bacteria plate prepared in step 1.9 using a sterile spatula. Incubate plates at 20 °C for 3-4 days. Under a stereo microscope, visually confirm the presence of gravid adults.
- Preparation of synchronized population of worms
- Collect gravid adults from at least 2 plates by dispensing autoclaved deionized water all around the plate using a squirt bottle. Gently swirl the plate to dislodge the worms and collect the worms into a 15 mL polystyrene conical tube using a disposable plastic pipette.
- Spin down the tube in a centrifuge at 140 x g for 2 min to pellet the worms, then aspirate off supernatant using a vacuum pump or with built in laboratory vacuum.
- Resuspend and wash the worms by filling the tube with autoclaved deionized water and mix and centrifuge at 140 x g for 2 min. Aspirate the supernatant and repeat this last step two more times or until worms are clear from bacteria (water appears clear when mixed with the worms).
- Add 5 mL of freshly made sodium hypochlorite/NaOH solution (step 1.3) to the worm pellet and rapidly mix using a vortex. Incubate the tube on a rocker for about 4-8 min. The time of incubation with sodium hypochlorite/NaOH solution fluctuates between 4-8 minutes based on the quality of the stock sodium hypochlorite solution (bleach).
- Put a drop (2-50 µL) of solution containing worms on a glass microscope slide and check every 2 min under the microscope for worm lysis. When about 70% of the worms are lysed and eggs are released, fill the tube with egg buffer prepared in step 1.2 and immediately centrifuge for 1 min at 140 x g to pellet the embryos and worm carcasses.
- Aspirate the supernatant and wash the pellet 3 more times by filling the tube each time with egg buffer. Spin down at 140 x g for 1 min and remove the supernatant each time. The pellet turns white at the end of washes.
- After the final wash, separate the embryos from the dead carcasses in 30% sucrose solution. Add 5 mL of autoclaved deionized water to the pellet, resuspend and add 5 mL of 60% sucrose prepared in step 1.4. Mix thoroughly and centrifuge at 160 x g for 6 min.
- Use a glass Pasteur pipette to transfer the embryos floating at the upper meniscus into a fresh 15 mL conical tube. Do not take more than 3-4 mL. To remove any remaining sucrose, wash the embryos 3 times with autoclaved water by centrifuging at 140 x g for 3 min, removing the supernatant and resuspending the pellet (and filling the tube) each time.
- Repeat the washes with 1x M9 buffer. After the final wash, resuspend the pellet in 10 mL of M9. Leave the tubes on a shaker overnight (no more than 14 h) for the eggs to hatch into L1 larvae. Worms will remain in L1 larval stage due to lack of food.
- Wash the L1 larvae 3 times with autoclaved water to remove any pheromones released by the larvae by centrifuging at 140 x g for 2 min. Resuspend the larvae in 1 mL of water. Make a 1:10 dilution of the worms in water, pipette a 10 µL drop on a glass slide, put a coverslip on and count the number of worms under a stereoscope. Repeat this twice and average the results.
- Pipette the volume of worms that corresponds to about 1,000 worms onto an NA22 plate (that was previously brought to room temperature) by placing small drops on the plate. Leave the plate half-open until the drop dries out. Then cover the plate and incubate upside down in 20 °C incubator for about 44-48 h or until the worms reach late L4 stage, as confirmed visually under a stereomicroscope. Now the worms are ready to be tested for SWIP.
3. SWIP
NOTE: We describe the manual method of assessing SWIP in wild-type worms treated with AMPH. We also briefly discuss the tracking of worms and further analysis of worm kinetics using an automated worm tracker and a tracking software which were previously described by Hardaway et al.10.
- Manual method to test for SWIP
- Aliquot 40 µL of 200 mOsm/L sucrose solution either with or without 0.5 mM AMPH into a glass spot plate. Under the stereoscope, pick 8-10 late-L4 stage worms with an eyelash or platinum pick and submerge the pick in the plate containing the solution until worms move off the pick and swim into the solution. Note the number of worms picked into the well, start the timer, observe and record the number of worms exhibiting SWIP at each minute mark.
- Copy the raw data into a spreadsheet and calculate the percent of worms paralyzed by dividing the number of worms paralyzed at each minute by total number of worms tested throughout the assay and multiply by 100. Copy the percent values into any graphing and statistical software and plot the data with percent values on the Y axis and time on X axis using the XY graph format.
- Perform two-way ANOVA followed by post-hoc analysis (e.g., Bonferroni post-test) to test for statistical significance among control, AMPH groups and time of treatment.
- Automated analysis of SWIP
- Perform automated analysis on a single worm at a time. The protocol to set up camera, the worm tracker software and script to run the tracking software analysis are described in detail in Hardaway et al.16.
- Briefly, place a single late L4 stage hermaphrodite into a glass spot plate utilizing an eyelash pick, as described in the manual method in section 3.1.2. Record swimming videos of one worm at the time and use the worm tracker software to calculate the frequency of body bends.
- Follow the script provided with the tracking software to obtain worm thrashing frequency and to generate heat maps from the worm thrashing data.
Representative Results
We present an example of SWIP assay induced by AMPH treatment. Figure 1 shows a schematic representation of the assay setup as described above. For the manual assay, about 8-10 age synchronized late L4 stage worms are collected with an eyelash or platinum pick and placed into a glass spot plate filled with 40 µL of 200 mOsm/L sucrose (control solution) or sucrose with 0.5 mM AMPH and tested for SWIP.
When animals stop swimming (i.e., exhibit SWIP), they quickly sink to the bottom of the well and do not move. Therefore, the discrimination between animals that still swim on the surface of the water versus the ones that are steady at the bottom of the well is very straightforward. Most of the worms tested in control solution swim continuously for at least 10 minutes, whereas under AMPH treatment, the number of animals exhibiting SWIP progressively increases. The maximal percentage of animals exhibiting SWIP is proportional to the concentration of AMPH used1,5,13. When DAT-1 knockout (dat-1) worms are tested in control solution, 40-70% worms exhibit SWIP within 10 minutes4,13. This result is comparable to the percentage of paralyzed animals measured in wild-type animals treated with 0.5 mM AMPH (Figure 2).
Worms exposed to either sucrose or sucrose containing AMPH do not show SWIP in the first minute of observation (Figure 2). However, while worms treated with sucrose continue to swim for 10 minutes, worms treated with AMPH start to exhibit SWIP after 2 minutes of treatment and after 10 minutes, 66 ± 3% animals show SWIP (Figure 2).
For the automated analysis, videos of worms under control or AMPH treatments are recorded one worm at a time using a video recording software. A computer tracking software is used to track worm thrashing and the resulting data are imported into and analyzed with the software suit. Samples of heat maps of animals exposed to control or AMPH, displaying actively moving animals in red and paralyzed worms in green, are shown in Figure 3.
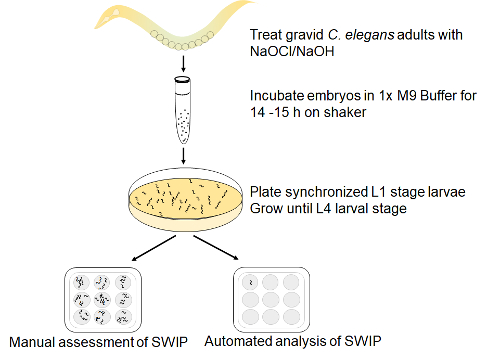
Figure 1: Assay set up for SWIP. Gravid adult wild-type (N2) worms were lysed with sodium hypochlorite/NaOH treatment to release embryos. The embryos were allowed to hatch and develop into synchronized L1 larvae in M9 buffer for 14 h on a shaker and then plated on an NGM plate seeded with NA22 bacteria. After 42-48 h late L4 stage larvae were visually identified under the stereoscope and picked with an eyelash pick into a spot plate with or without amphetamine in control sucrose solution and scored for SWIP either manually or through automated analysis. Please click here to view a larger version of this figure.
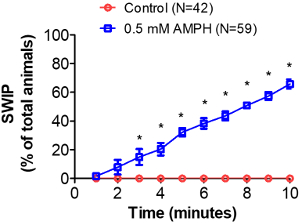
Figure 2: Amphetamine-induced SWIP using manual assay. Worms in sucrose or sucrose with 0.5 mM amphetamine (AMPH) were visually scored for SWIP behavior every minute using a stereoscope. The percent of animals exhibiting SWIP was calculated by dividing the number of paralyzed worms by the total number of worms assayed for each time point, and then multiplying the result by 100. The percent of worms exposed to AMPH (blue squares) showing SWIP increases overtime, while the untreated worms (red circles) continue to swim during the 10-minute window. N represents the number of animals tested in each group. Error bars indicate standard error of means (SEM). Statistical significance was assessed by performing two-way ANOVA with Bonferroni multiple comparison test (p < 0.0001). Please click here to view a larger version of this figure.
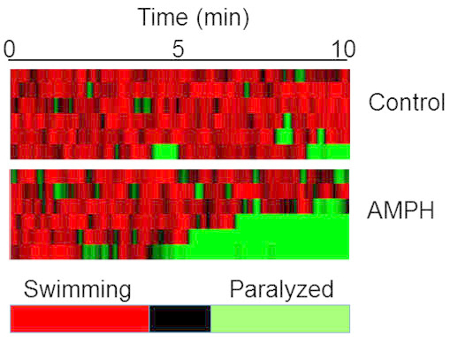
Figure 3: Amphetamine-induced SWIP via automated analysis. Videos of worms in sucrose or sucrose with 0.5 mM AMPH were recorded using a camera mounted on a stereoscope. Swimming videos of individual worms were tracked with a tracking software and analyzed using tracking software suit. Heat maps were generated from the data where red areas show the worms that are actively moving, and green areas indicate paralyzed worms. Each experimental group is representative of 6 animals. Please click here to view a larger version of this figure.
Discussion
Here, we describe a step-by-step protocol to perform a behavioral assay, SWIP, in C. elegans. This protocol is simple and straightforward with no major technical hurdles making this assay very user friendly. Nevertheless, there are some critical aspects that need to be considered in order to effectively perform the assay.
Care should be taken to ensure that the worms used for the assay are well fed, since dietary restriction affects SWIP17. Gentle handling of worms while picking as well as timed sodium hypochlorite/NaOH treatment during lysis are critical steps, as trauma during picking (more common when using a platinum pick) or extended exposure of embryos to sodium hypochlorite/NaOH solution can cause permanent damage to the worms18 and thus compromise their ability to swim.
Sterile techniques should be followed to avoid contamination. Contamination of the agar plates compromises the health of the animals and thus alters their ability to swim. Slight difference in the percentage of animals exhibiting SWIP can be observed using agar plates seeded with different strains of E. coli bacteria (NA22, OP50, etc.). In our protocol, we use NA22 to yield a large number of worms.
Glass-spot plates, used during the SWIP assay, are preferred to plastic plates because they can be thoroughly washed, autoclaved and re-used when different types of drugs are tested.
Another important factor to be considered in a SWIP assay is the osmolarity of the liquid media in which the animals are tested19. In our protocol, we use sucrose to bring the osmolarity of water up to 200 mOsm/L which was previously shown to be an optimal condition for the animals (Blakely RD. personal communication). Water with a controlled osmolarity is preferred because it eliminates possible differences in water quality over multiple assays performed in different days, weeks or months. Notably, dat-1 and wild-type animals treated with AMPH do not exhibit SWIP if salty solutions (e.g., M9 solution) are used as control media.
The time required for most of the animals to exhibit SWIP can slightly change among different worm stages (L1-L4). For example, Masoudi et al. (2014) reported that after 5 min 80% of dat-1 L1 animals still swim, thus only 20% of animals exhibit SWIP. On the other hand, only 50% of L4 dat-1 animals still swim after 5 min20. Therefore, it is important that animals tested for SWIP are assayed at the same age. We have optimized our assay using always late L4 staged animals. Late L4 larvae have the advantage of being easily recognizable among the other larval stages because, at this stage, animals have reached their adult size and exhibit a characteristic thin line dividing the white spot in the center of their body that will later differentiate into a mature vulva. The time-span of the assay is also critical. For example, after 15 minutes 90% of L4 dat-1 mutants exhibit SWIP but at later time (30 min), only 60% of them exhibit SWIP20.
The automated SWIP assay eliminates human errors and improves high-throughput screening with respect to manual assays. However, tracking software programs are time consuming since they can only track a single worm at a time.
With respect to other C. elegans DA-dependent behaviors, SWIP is a less time-consuming type of assay. For instance, the basal slowing response2 is not an immediate-type of assay. In fact, worms need to be chronically fed with the drugs, and this could result in penetrant and off-target effects. Thus, it might not be as effective to screen for drugs.
One of the major applications of SWIP is to screen for various drugs that target the dopaminergic pathway. In cases where the drugs are not water soluble (e.g., mazindol), proper dilutions should be performed to achieve concentrations where the carrier solution is not toxic to the worms. Moreover, if different concentrations of the same drug are used5,13,14, dose-response curves can also be analyzed. For example, the rate of progression (slope of the curve of animals per minute, Figure 2) can be reported as function of the concentration and used to compare the effects of different drugs (e.g., AMPH vs cocaine).
When SWIP is used to investigate the mechanism of action of drugs at the dopaminergic synapses, attention should be paid if results are extended to other animals. For instance, imipramine, a specific inhibitor of the mammalian norepinephrine transporter (NET) has been used to induce DA-mediate SWIP in N2 animals4. C. elegans does not synthetize norepinephrine and consequently does not express NET. However, the C. elegans DA transporter shares homology with mammalian NET21. For this reason, drugs that are specific inhibitors of mammalian NET and have limited effects on mammalian DAT (e.g., imipramine) show high selectivity to C. elegans DAT. Thus, species selectivity might limit our ability to extrapolate findings from C. elegans to humans.
The most important factor to consider when designing SWIP assays is the inclusion of experiments proving that SWIP is mediated by the dopaminergic system. In fact, impaired swimming could be generated by factors other than genes related to the DA system (e.g., general defects in muscles contraction). To ensure that SWIP is indeed mediated by DA, protocols should include experiments performed with animals in which DA has been depleted. This can be achieved by either using knockout animals lacking expression of cat-2, the C. elegans homologue of the tyrosine hydroxylase, which is the rate-limiting enzyme for DA synthesis, or by pre-treating wild-type animals with reserpine, a drug that causes DA depletion from vesicles3. For instance, McDonald et al.3 showed that basal SWIP observed in dat-1 mutants was recovered when these animals were pre-treated with reserpine. This result suggests that SWIP is DA-mediated. On the other hand, using cat-2, dat-1 and mutants lacking expression of each of the dopaminergic receptors, Safratowich et al. (2014) demonstrated that the trace amine β-phenylethylamine (βPEA) induces SWIP within 1 minute of treatment independently from DA but by direct activation of the ligand-gated ion channel LGC-5514. The authors showed that βPEA- and DA-induced SWIP are mechanistically different and they can be easily discriminated experimentally. In fact, while βPEA-induced SWIP is DA- and dop-3-independent, it reaches maximal values within 1 min and rapidly decreases after 2 min14, DA-mediated SWIP is cat-2 and dop-3 dependent and is essentially zero after 1 minute (Figure 2). Thus, there is a large difference in the time required to reach maximal effects, and this allows to quickly discriminate between the two phenomena: 1) the fast βPEA-induced SWIP within 1 minute obtained by direct activation of the LGC-55 channels and 2) the slow DA-mediated SWIP which occurs when a surplus of extracellular DA builds up over time (10-15 min) and the DA receptors DOP-3 are overstimulated3.
In conclusion, with the right set of experiments, which include the use of knockout animals for key player genes of the dopaminergic system (cat-2, dat-1, dop-3) and the use of drugs depleting DA storages (reserpine), SWIP has been successfully used to elucidate the mechanism of action of drugs like AMPH5,13, βPEA14,15 and azaperone6.
Disclosures
The authors have nothing to disclose.
Acknowledgements
The authors would like to thank Dr. Osama Refai from Dr. Randy Blakely's lab for guidance with the automated analysis of SWIP. This work was supported by funding from NIH R01 DA042156 to LC.
Materials
| Aluminum foil | Reynolds wrap | 1091835 | |
| Amphetamine | Sigma | 51-63-8 | |
| Autoclave | |||
| Bacterial Incubator | New Brunswick scientific | M1352-0000 | |
| Bacteriological grade, Agar | Lab Scientific, Inc | A466 | |
| Bacto (TM) Peptone | BD | REF 211677 | |
| Calcium Chloride (dihydrate) | Sigma-Aldrich | C3881 | |
| Camera | Thorlabs | U-CMAD3 | |
| Centrifuge | Eppendorf 5810R 15amp | E215059 | |
| Cholesterol | Sigma-Aldrich | 57-88-5 | |
| Deionised water | Millipore | Z00QSV0WW | Milli-Q |
| Depression glass spot plate | Corning | Corning, Inc. 722085 | |
| Erlenmeyer flask | ThermoFisher | 4103-0250PK | |
| Eye lash | |||
| Glass slide | Fisherbrand | 12-550-15 | |
| Graphing and statistical software | Prism | Graphpad 5 | |
| HEPES | Sigma-Aldrich | RB=H3375 & H7006 | |
| Hypochlorite | Hawkins | Sodium Hypochlorite 4-6%, USP" 1 gal | |
| LB Broth, Miller | Fisher | BP1426 | |
| Magnesium Chloride (Hexahydrate) | Sigma-Aldrich | RB=M0250 | 500g |
| Magnesium sulfate (heptahydrate) | Sigma-Aldrich | M1880 | |
| Magnetic stir bar | Fisherbrand | 16-800-510 | |
| Microcentrifuge tubes | ThermoFisher | 69715 | |
| NA 22 bacteria | CGC | ||
| Nystatin | Sigma | 1400-61-9 | |
| Osmometer | Advanced Instruments, Inc | Model 3320 | |
| Pasteur Pipettes | Fisherbrand | 13-678-20A | |
| Petriplates | Falcon | 351007 | |
| pH Meter | Orion VersaStar Pro | IS-68X591202-B 0514 | |
| Polystrine conical tubes | Falcon | 352095 | |
| Potassium Chloride | Sigma-Aldrich | P9541 | |
| Potassium dihydrogen phosphate | Sigma-Aldrich | 7778-77-0 | |
| Potassium Phosphate – DIBASIC | Sigma-Aldrich | P-8281 | |
| Potassium Phosphate – MONOBASIC | Sigma-Aldrich | P0662 | |
| Serological pipettes | VWR | 10ml=89130-898 | |
| Shaker | Reliable Scientific | 55S 12×16 | |
| Sodium Chloride | Fisher | RB=BP358-1 | |
| Sodium dihydrogen Phosphate | Fisher | RB=S381 | |
| Spreadsheet | MS office | Microsoft Excel | |
| Stereo Microscope | Zeiss | Model tlb3. 1 stemi2000 | |
| Sterile Pipette tips | Various | 02-707-400 | |
| Sucrose | Sigma-Aldrich | RB=S5016 | |
| Superglue | Loctite | 1647358 .14 oz. | |
| SwimR sofware | 10.18129/B9.bioc.SwimR | ||
| Tracker 2 | Worm Tracker 2.0 | www.mrc-lmb.cam.ac.uk/wormtracker/ | |
| Video recording software | Virtualdub | http://www.virtualdub.org/ |
References
- de Bono, M., Villu Maricq, A. Neuronal Substrates of Complex Behaviors in C. elegans. Annual Review of Neuroscience. 28 (1), 451-501 (2005).
- Sawin, E. R., Ranganathan, R., Horvitz, H. R. C. elegans Locomotory Rate Is Modulated by the Environment through a Dopaminergic Pathway and by Experience through a Serotonergic Pathway. Neuron. 26 (3), 619-631 (2000).
- McDonald, P. W., et al. Vigorous Motor Activity in Caenorhabditis elegans Requires Efficient Clearance of Dopamine Mediated by Synaptic Localization of the Dopamine Transporter DAT-1. Journal of Neuroscience. 27 (51), 14216-14227 (2007).
- Carvelli, L., Blakely, R. D., DeFelice, L. J. Dopamine Transporter/Syntaxin 1A Interactions Regulate Transporter Channel Activity and Dopaminergic Synaptic Transmission. Proceedings of the National Academy of Sciences of the United States of America. 105 (37), 14192 (2008).
- Carvelli, L., Matthies, D. S., Galli, A. Molecular mechanisms of amphetamine actions in Caenorhabditis elegans. Molecular Pharmacology. 78 (1), 151-156 (2010).
- Refai, O., Blakely, R. D. Blockade and reversal of swimming-induced paralysis in C. elegans by the antipsychotic and D2-type dopamine receptor antagonist azaperone. Neurochemistry International. , (2018).
- Bermingham, D. P., et al. The Atypical MAP Kinase SWIP-13/ERK8 Regulates Dopamine Transporters through a Rho-Dependent Mechanism. The Journal of Neuroscience. 37 (38), 9288-9304 (2017).
- Nass, R., et al. A genetic screen in Caenorhabditis elegans for dopamine neuron insensitivity to 6-hydroxydopamine identifies dopamine transporter mutants impacting transporter biosynthesis and trafficking. Journal of Neurochemistry. 94 (3), 774-785 (2005).
- Hardaway, J. A., et al. Forward genetic analysis to identify determinants of dopamine signaling in Caenorhabditis elegans using swimming-induced paralysis. G3. 2 (8), 961-975 (2012).
- Hardaway, J. A., et al. Glial Expression of the Caenorhabditis elegans Gene swip-10 Supports Glutamate Dependent Control of Extrasynaptic Dopamine Signaling. Journal of Neuroscience. 35 (25), 9409-9423 (2015).
- Felton, C. M., Johnson, C. M. Dopamine signaling in C. elegans is mediated in part by HLH-17-dependent regulation of extracellular dopamine levels. G3. 4 (6), 1081-1089 (2014).
- Lanzo, A., et al. Silencing of Syntaxin 1A in the Dopaminergic Neurons Decreases the Activity of the Dopamine Transporter and Prevents Amphetamine-Induced Behaviors in C. elegans. Frontiers in Physiology. 9 (576), (2018).
- Safratowich, B. D., Lor, C., Bianchi, L., Carvelli, L. Amphetamine activates an amine-gated chloride channel to generate behavioral effects in Caenorhabditis elegans. The Journal of Biological Chemistry. 288 (30), 21630-21637 (2013).
- Safratowich, B. D., Hossain, M., Bianchi, L., Carvelli, L. Amphetamine Potentiates the Effects of -Phenylethylamine through Activation of an Amine-Gated Chloride Channel. Journal of Neuroscience. 34 (13), 4686-4691 (2014).
- Carvelli, L. Amphetamine activates / potentiates a ligand-gated ion channel. Channels (Austin). 8 (4), 294-295 (2014).
- Hardaway, J. A., et al. et al.An open-source analytical platform for analysis of C. elegans swimming-induced paralysis. Journal of Neuroscience Methods. 232, 58-62 (2014).
- Lüersen, K., Faust, U., Gottschling, D. -. C., Döring, F. Gait-specific adaptation of locomotor activity in response to dietary restriction in Caenorhabditis elegans. The Journal of Experimental Biology. 217, 2480-2488 (2014).
- Porta-de-la-Riva, M., Fontrodona, L., Villanueva, A., Cerón, J. Basic Caenorhabditis elegans methods: synchronization and observation. Journal of Visualized Experiments. (64), e4019 (2012).
- Lamitina, S. T., Morrison, R., Moeckel, G. W., Strange, K. Adaptation of the nematode Caenorhabditis elegans. to extreme osmotic stress. American Journal of Physiology-Cell Physiology. 286 (4), 785-791 (2004).
- Masoudi, N., Ibanez-Cruceyra, P., Offenburger, S. -. L., Holmes, A., Gartner, A. Tetraspanin (TSP-17) Protects Dopaminergic Neurons against 6-OHDA-Induced Neurodegeneration in C. elegans. PLoS Genetics. 10 (12), 1004767 (2014).
- Jayanthi, L. D., et al. The Caenorhabditis elegans gene T23G5.5 encodes an antidepressant- and cocaine-sensitive dopamine transporter. Molecular Pharmacology. 54 (4), 601-609 (1998).
