We have demonstrated how the procedures described above can be used to relatively quantify the cytotoxicity of S. aureus against human PMNs using MRSA PFGE-type USA300 and an isogenic deletion mutant of saeR/S in this strain (USA300ΔsaeR/S) generated in previous studies6. PMNs isolated using the procedures described in section 1 of this protocol were stained with propidium iodide and examined using flow cytometry. Forward and side scatter plots were used to illustrate contamination of purified PMNs by monocytes or lymphocytes (Figure 1A,B) and PMN integrity was determined using propidium iodide staining (Figure 1C). The described method of human PMN purification can consistently yield 0.5 x 107 to 1 x 108 PMNs that are >98% pure and are >95% propidium iodide negative.
The cytotoxicity of extracellular proteins produced by USA300 and USA300ΔsaeR/S were tested against purified PMNs (Figure 2) following the procedures described in section 2 of this protocol. These experiments demonstrate a concentration dependent increase in the propidium iodide staining of purified PMNs following 30 min of intoxication with extracellular proteins produced by USA300 (Figure 2B). Previous studies have demonstrated that the SaeR/S two-component system is important for expression of numerous bi-component leukocidins that target human PMNs6,10,11,16. Congruent with these previous findings, very few propidium iodide-positive PMNs were detected following exposure to extracellular proteins produced by USA300ΔsaeR/S (Figure 2B). Further experiments demonstrated a steady increase in the proportion of lysed PMNs following intoxication by USA300 extracellular proteins that plateaued after approximately 30 min (Figure 2C). Minimal lysis of human PMNs was noted at all timepoints following exposure to extracellular proteins produced by USA300ΔsaeR/S. These results illustrate the utility of this assay for the relative quantification of cytotoxicity by extracellular S. aureus proteins against human PMNs.
We tested USA300 and USA300ΔsaeR/S using the S. aureus cytotoxicity assay against human PMNs following phagocytosis that is described in section 3 of this protocol (Figure 3). A concentration dependent increase in the proportion of propidium iodide positive PMNs was observed 90 min after the phagocytosis of USA300 (Figure 3A). A significant decrease was observed in the proportion of PMNs that were propidium iodide positive following the phagocytosis of USA300ΔsaeR/S (Figure 3A), supporting other results that indicate the SaeR/S two-component system is important for the cytotoxicity of S. aureus against human PMNs (Figure 2)7,11. As previously mentioned and demonstrated in Figure 3A, differences in S. aureus concentration have a pronounced impact on PMN lysis following phagocytosis. Enumeration of the USA300 and USA300ΔsaeR/S inoculum used in each of these experiments demonstrated that the contrast in cytotoxicity between these strains was not due to differences in the concentration of bacteria used (Figure 3B). These findings show how the S. aureus cytotoxicity assay against human PMNs following phagocytosis can be used to assess the ability of different S. aureus strains to compromise human PMN plasma membrane integrity.
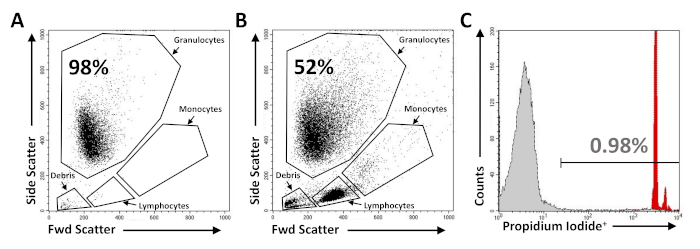
Figure 1: Flow cytometry analysis of purified PMNs. Representative flow cytometry dot plots of (A) purified human PMNs and (B) PMNs that have been purposely contaminated with peripheral blood mononuclear cells. (C) Representative flow cytometry histogram demonstrating minimal propidium iodide staining (<1%) of purified PMNs (shaded grey) as compared to PMNs treated with 0.05% Triton X-100 (shaded red). Please click here to view a larger version of this figure.
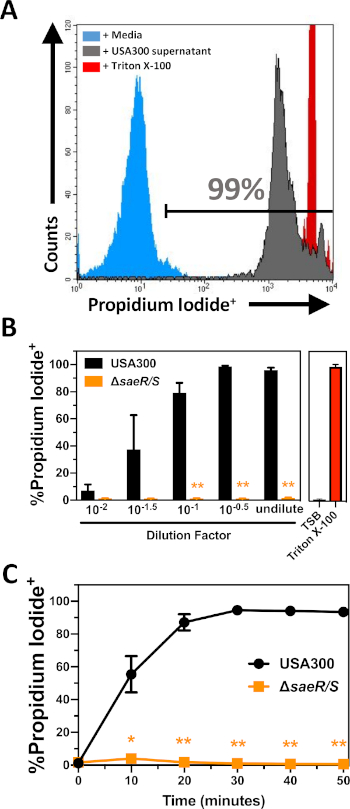
Figure 2: Flow cytometry analysis of PMNs intoxicated with extracellular proteins produced by S. aureus. (A) Representative flow cytometry histogram of PMNs stained with propidium iodide after 30 min of incubation with media control (shaded blue), filtered USA300 supernatant at a final concentration of 1:110 (shaded grey), or 0.05% Triton X-100 (shaded red). (B) The proportion of propidium iodide positive PMNs after 30 min of incubation with different concentrations of USA300 or USA300∆saeR/S supernatants. (C) The proportion of propidium iodide positive PMNs over time following incubation with USA300 or USA300∆saeR/S supernatant at a final concentration of 1:110. Data are presented as mean ± SEM of at least 3 separate experiments with * p ≤ 0.05 and ** p ≤ 0.005 as determined by two-tailed t-test. Please click here to view a larger version of this figure.
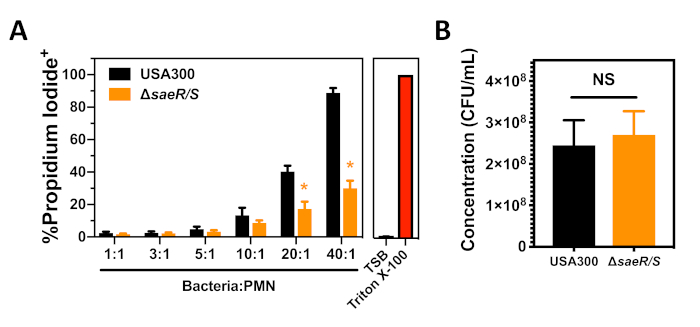
Figure 3: Flow cytometry analysis of PMNs following phagocytosis of S. aureus. (A) The proportion of propidium iodide positive PMNs 90 min after the phagocytosis of different concentrations of USA300 or USA300∆saeR/S. (B) Concentration of opsonized S. aureus strains used for the experiments shown in panel A. Data are presented as mean ± SEM of 4 separate experiments with * p ≤ 0.01 as determined by two-tailed t-test. Please click here to view a larger version of this figure.
