Wild type Nipponbare (Figure 2A) and ssg1 sections (Figure 2B) were examined under three magnifications: 260x, 920x, and 4200x. This technique allows for the preparation of sections of sufficient quality to observe the entire endosperm cell (Figure 3A), compound starch granules (Figure 3B), and individual subgranules (Figure 3C). Husked kernels take longer to process than polished kernels as the dry hulls need to be removed by abrasion before sectioning. Chalky kernels also take longer to process than polished translucent kernels, as care must be taken to not shatter the kernel during sectioning. A properly prepared rice section should be approximately 0.9 mm thick (Table 1) with minimal to no shattering of the endosperm (Figure 1N) and intact pericarp and aleurone layers (Figure 1O). Improper placement of the scalpel on the pipette tip when sectioning can lead to ‘chipped’ sections (Figure 1P). Similarly, bright field images of optimal transverse sections of ssg1 (Figure 1Q) demonstrated intact endosperm, pericarp, and aleurone layers intact and available for visualization (Figure 1R). A broken chalky kernel section (Figure 1S) may still be useable for visualization if the only purpose is to observe SGs, but endosperm cell pattern will not be visible. A broken section may be difficult to handle for analysis. More shearing of endosperm cell walls was observed in wild type Nipponbare, as the cells are more tightly packed and less friable than the ssg1 kernels. No shearing of endosperm cells was observed in the ssg1 sections and compound starch granules are intact.
Figure S1 demonstrates the reliability of results using the ‘telescope’ technique to section rice kernels. Rice lines identified as translucent kernel producers – wild type Resistant Starch (RS) hybrid line Xieyou 7954 (Oryza sativa L. ssp. indica)12,13,14 (Figure S1A) and cobalt-generated mutant RS11113,15 (Figure S1B) produced sections through which light was visible using a stereomicroscope. The corresponding SEM images revealed that these lines produce the ‘normal’ rice endosperm phenotype: tightly packed, polyhedral starch granules. Chalky kernel producers, commercial variety Yi-Tang16 (Figure S1C) and RS413, a mutant of RS11115 (Figure S1D), exhibited white, opaque kernel sections. The corresponding SEM images displayed markedly different morphology compared to the wild type translucent RS background line: starch granules were round and loosely packed. Wild type Xiushui 11 (Oryza sativa L. ssp. japonica) (Figure S1E) and its mutant, KMD1 (Kemingdao1), which express the Cry1Ab gene to inhibit insect predation17,18,19 (Figure S1F) exhibited sections and endosperm morphotypes similar to the translucent RS lines.
The technique presented here is optimal for preparing samples of chalky-type rice kernels for phenotypic analysis, but also provides advantages for sectioning translucent rice kernel phenotypes20: slicing the samples using pressure from above reduces risk of shattering of the endosperm and dislocation. Samples can easily be prepared within seconds (Table 2). Multiple genotypes were analyzed using this technique to test its efficacy (Table 3). As shown in Figure S2, this technique can be applied to seeds of other species. The model monocot Brachypodium distachyon produces very hard seeds containing only B-granule starch21, which lack puroindoline A, a protein that confers softness to starch granules22. It was still possible to obtain an intact transverse section (Figure S2A). Obtaining an intact transverse section from soft white winter wheat (SWWW) was challenging but can be performed (Figure S2B). SWWW seeds are high in puroindoline A and large when compared to B. distachyon seeds and rice kernels. These seeds crumble frequently when sectioning using the telescope assemblage.
| Genotype | Mean section width (µm) using telescope assemblage | Mean section width (µm) sectioning freehand |
| Nipponbare (husked) | 971.7 ± 152.4ab | 1059.571 ± 394.2ab |
| Xieyou 7954 | 825.1 ± 128.3b | 1306.187 ± 179.1a |
| RS4 | 910.6 ± 165.0ab | 1126.694 ± 395.3ab |
| Means followed by the same letters are not significantly different at P < 0.01 using a one-way analysis of variance (ANOVA) and Tukey’s test (n = 10). Statistical analyses were performed using JMP 15 software. | ||
Table 1: Mean kernel section thickness.
| Genotype | Mean time (s)* |
| Nipponbare (husked) | 14.7 ± 1.36a |
| Xieyou 7954 | 9.81 ± 0.98b |
| RS4 | 11.9 ± 1.28c |
| *Using the telescope assemblage. | |
| Means followed by the same letters are not significantly different at P < 0.01 using a one-way analysis of variance (ANOVA) and Tukey’s test (n = 10). Statistical analyses were performed using JMP 15 software. | |
Table 2: Mean sample preparation time.
| Genotype | Background | Quality |
| Nipponbare | Wild type | Translucent |
| Substandard starch grain1 (ssg1) | Nipponbare | Chalky |
| Resistant Starch (RS) Xieyou 7954 | Wild type | Translucent |
| RS111 | Xieyou 7954 | Translucent |
| RS4 | RS111 | Chalky |
| Yi-Tang, ‘New Life’, Lujuren brand | Xieyou 7954 | Chalky |
| Xiushui 11 | Wild type | Translucent |
| Kemingdao1 (KMD1) | Xiushui 11 | Translucent |
Table 3: Rice genotypes examined in this study.

Figure 1: Preparation of transverse rice sections. (A) Wild type Nipponbare kernel with intact husk. (B). Kernel placed on a flat four-inch diameter rubber stopper. (C) Husks were removed by grinding the kernel between two apposing rubber stoppers. (D) Husk has been separated from the rice kernel. (E) Close-up of husked rice kernel. Embryo end is indicated. (F) Insertion of kernel into pipette tip using fine forceps. (G) Kernel was lodged into the distal end of the pipette tip. (H) Insertion of the second pipette tip to immobilize the kernel for sectioning (the ‘telescope’ assemblage). (I) The rice kernel was fitted snugly into the distal end of the pipette tip. (J) Sectioning of the rice kernel within the assemblage. (K) Close-up of the section cut. (L) A section of the kernel enclosed by the plastic annulus. (M) Close-up of the transverse section. (N) Transverse section of wild type Nipponbare. (O) Close-up of the endosperm within the wild type Nipponbare section. (P) Poor, suboptimal section of wild type Nipponbare kernel. (Q) Transverse section of Nipponbare mutant ssg14. (R) Close-up of the endosperm within the ssg1 section. (S) Poor, suboptimal section of ssg1. Bar (panels A, N–S) = 1 mm. Whole rice kernel and sections were imaged using a stereomicroscope with a digital zoom camera and gooseneck lights. Please click here to view a larger version of this figure.
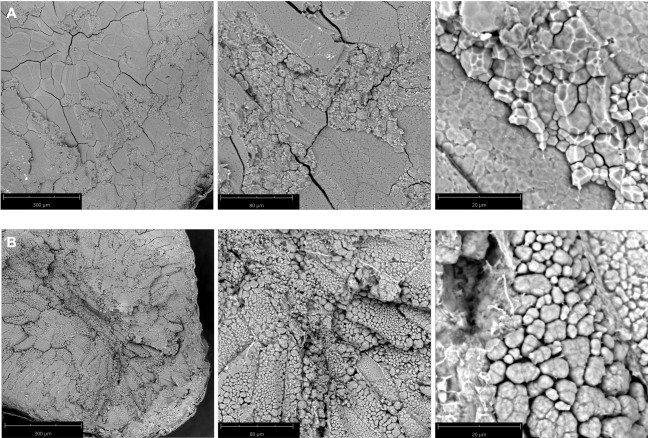
Figure 2: SEM images of transverse kernel sections. (A) Wild type Nipponbare, a translucent cultivar. The compound starch granules were cemented tightly to one another; (B) Nipponbare mutant ssg14, a chalky phenotype. The compound starch granules were loosely packed and lack the cementitious nature of wild type Nipponbare starch morphotype. Magnification from left to right: 260x, 920x, and 4200x. Bar length is indicated in panels. Please click here to view a larger version of this figure.
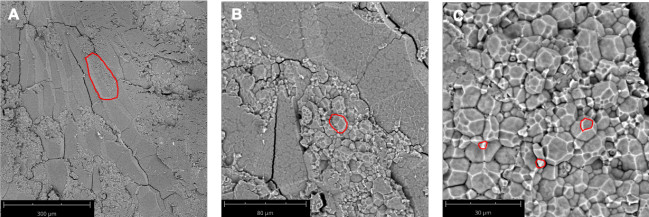
Figure 3: SEM microscopic anatomy of a transverse kernel section of Xiushui 11. (A) A single endosperm cell is outlined in red. 260x magnification. (B) A compound starch granule is outlined in red. 920x magnification. (C) Multiple starch subgranules are outlined in red. 2250x magnification. Bar lengths are indicated in the panels. Please click here to view a larger version of this figure.
Figure S1: Transverse sections of other rice genotypes prepared for SEM using this technique. (A) Resistant Starch (RS) Xieyou 795412. (B) RS111, a high-RS transparent mutant of 795413. (C) RS4, a chalky mutant of RS11115. (D) Yi-Tang, a commercial variety of high amylose rice16. (E) Xiushui 11. (F) KMD1 (Kemingdao1)17,18,19. 10x magnification for bright field images. White bar = 1 mm. 2250x magnification for SEM images. Bar lengths are indicated in the panels. Please click here to download this figure.
Figure S2: Technique is useful for other seeds. (A) Transverse section of false purple brome (Brachypodium distachyon L. accession Bd21) seed. (B) Transverse section of soft white winter wheat (Triticum aestivum L. cv. Augusta) seed. Bright field, 20x magnification. Bar = 1 mm. Please click here to download this figure.
