The determining factor for the success of the procedure is preventing crypt contamination. Organoid development from the villi (and not from any contaminating crypts) is ensured by confirming four major criteria: 1) ensuring the purity of the harvested villi by microscopic examination before and after plating the villi in BME-R1, 2) plating limited number of villi per well to allow visualization of all the plated villi individually, 3) onitoring the development of organoid daily; images of the time course show development of organoid from villi (Figure 3), and 4) onitoring the kinetics and morphological appearance of the organoids initiating from villi; the organoids initiating from villi appear irregularly shaped at first and take two to five days before they can be seen under 4x magnification as opposed to the crypt-derived organoids (cultured by isolating crypts using the EDTA/PBS chelation method1,4) that appear overnight as spherical structures with well-defined borders (Figure 2).
The optimal time for sacrificing the mice for testing the organoid-forming potential of the villi is after the dedifferentiating villi-epithelium express stem cell markers and begin the development of ectopic crypts in the villi in vivo (around 10 days after Tamoxifen injection of the Smad4f/f; Catnblox(ex3)/+; Villin-CreERT2 mice). These mice have a tamoxifen inducible cre-recombinase that causes Smad4 loss concomitant with β-catenin activation with tamoxifen. The ectopic crypts develop fully within two weeks of induction of the mutation, while the stem cells in the villus epithelium appear within a week after the induction of mutation in vivo. The number of villi that form organoids when plated in Matrigel has been reported to vary between 2% to 12%3. Harvesting the mutant mouse for plating the villi around the time when stem cells are present (around a week after the cre-recombinase induction) will prolong the time required for villi-derived organoids appear, whereas delaying the harvest to later than ten days after the cre-induction increases the risk crypt of contamination.
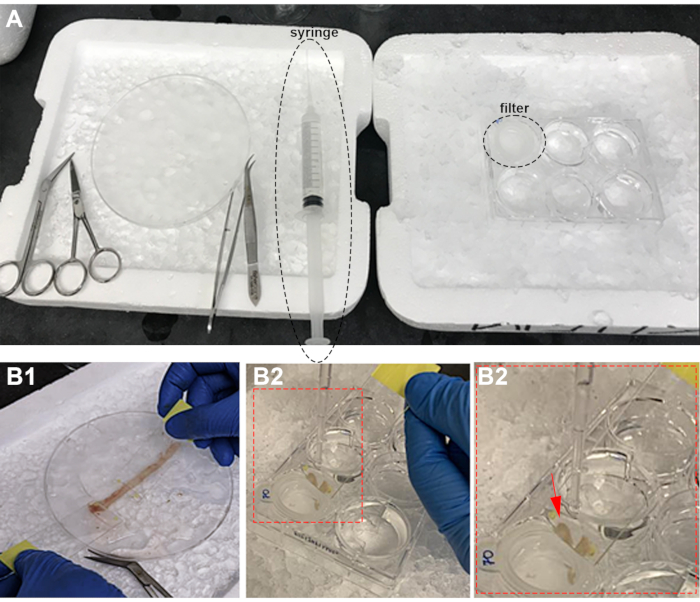
Figure 1: Experimental set-up and villi scraping from intestinal epithelium. A) PBS-filled syringe (syrg) with tubing and filter (Fltr) in PBS. B) Villi isolation by scraping (B1) and transferring to a filter (B2). The arrow points the villi collected on the slide. Please click here to view a larger version of this figure.
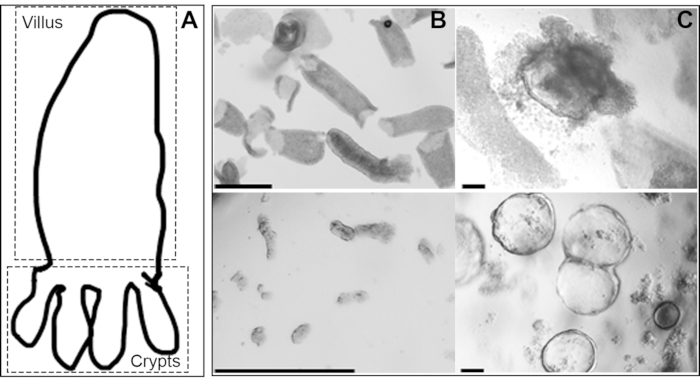
Figure 2: Distinction in the appearance of the organoids emerging from the crypts versus the dedifferentiated villi epithelium from the same mouse intestine: A) Cartoon showing villus and crypts. B) Whole mount (in PBS) of villi prepared by scraping (top panel) and crypts prepared by EDTA-chelation (lower panel). C) Villi-derived (top panel) and crypt-derived (lower panel) organoids from the same mouse intestinal epithelium in BME-R1 (scale bars, 500 um). Please click here to view a larger version of this figure.
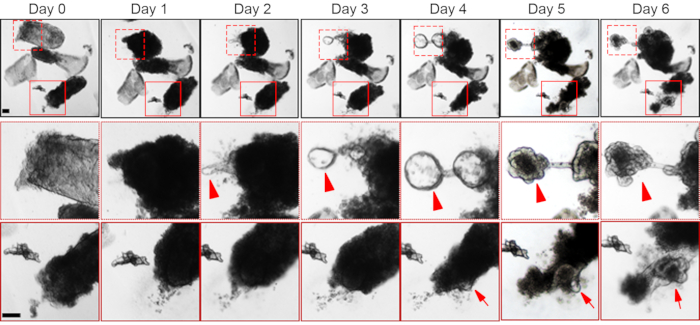
Figure 3: Time course of organoid formation from the dedifferentiating villi. Organoid initiation from two different villi is shown (the boxed regions are enlarged in the middle and bottom panels). The villi with organoid-forming potential appears dense, possibly due to the retention of the underlying mesenchyme. Organoid initiation from the villus is apparent at day two (solid arrow) from one of the villi (box with broken boundary), while in the other villus (box with solid boundary) the organoid appears at day four (arrow). The upper box in the day 6 panel shows a developed villi-derived organoid. Scale bar, 100 µm. Please click here to view a larger version of this figure.
