We developed customized mechanical probes, using nitinol filaments (Figure 1A,N), to elicit mechanically-evoked behaviors and generated a full behavioral dose response curve using both innocuous and noxious mechanical probes of varying intensity (Figure 2D) demonstrating that these probes can be used to study baseline (in the absence of injury) mechanical nociception.
Our behavioral assay results determined that probes exerting pressures below 200 kPa (~1.57 mN) (Figure 1M), when applied to Drosophila larvae, do not provoke an aversive rolling response (Figure 2D and Video 3). As expected, these subthreshold or non-noxious mechanical probes (175 kPa or 200 kPa) did not elicit visible neuronal tissue damage (Figure 2E). Because they do not induce damage, such probes could be useful to assess mechanical allodynia (hypersensitivity to a normally non-noxious mechanical stimuli). Conversely, suprathreshold or noxious probes (from 462 kPa to 5,116 kPa), elicited an augmented behavioral response (Figure 2D) in a dose dependent manner—with the higher pressures eliciting stronger behavioral responses. As anticipated, suprathreshold mechanical pressure also induced dose-dependent tissue damage to the peripheral sensory neurons themselves (Figure 2E). The measured area of tissue damage (in µm2 ± standard deviation) taken from four larvae for each group were: 2,051.03 ± 703.81 (462 kPa), 5,102.29 ± 1,004.67 (2,283 kPa), and 12,238.83 ± 3,724.11 (5,116 kPa). Thus, pressures greater than or equal to 462 kPa (~63 mN), which evoke an aversive rolling response (in 25% or more of the larvae) and cause visible neuronal tissue damage (Figure 2E), could be appropriate to study mechanical hyperalgesia (hypersensitivity to normally noxious mechanical stimuli). Nociceptive mechanical probes (≥462 kPa) always induce tissue damage (n = 10, evaluated qualitatively) but do not always provoke an aversive rolling response.
To evaluate mechanical hypersensitivity (allodynia and hyperalgesia), we used a well-established Drosophila larval model of nociceptive sensitization that uses ultraviolet light (UV) irradiation to induce tissue damage7,12. This assay has helped to dissect the genetic and cellular mechanisms of thermal nociceptive hypersensitivity8,9,10,13,14,15. To determine whether UV treatment causes mechanical allodynia, mid third-instar control (w1118) larvae were mock-irradiated or UV-irradiated (15–20 mJ/cm2) (Figure 3A). Then, the larvae were tested behaviorally at 2 h, 4 h, 8 h, 16 h, and 24 h post-treatment with a normally subthreshold mechanical probe (200 kPa, 1.57 mN). Approximately 20% of larvae responded as early as 2 h after UV treatment while 50% responded at 4 h, compared to 6.6% and 8.3% mock UV-irradiated animals, respectively (Figure 3B). This indicates that UV-induced tissue damage causes mechanical allodynia at 4 h post-irradiation. At later time points (8 h, 16 h, and 24 h) the behavioral response of the UV-treated larvae was in the range of 16%–20% responders (average mean of n = 3–6 sets of 10 larvae each), slightly increased (but not statistically significant) compared to the mock-irradiated control group (in the range of 3%–6% of responders, average mean of n = 3–6 sets of 10 larvae each) (Figure 3B).
To investigate mechanical hyperalgesia, a suprathreshold pressure (462 kPa, 3.63 mN), that normally induces an aversive rolling response in ~20% of larvae (Figure 2D) and causes neuronal tissue damage (Figure 2E), was used. We applied the 462 kPa probe onto the dorsal side of larvae with or without UV-induced tissue damage (Figure 3A). We found that larvae probed at 4 h, 8 h, and 16 h following UV treatment showed a significant increase in the aversive rolling response, with 4 h being the peak of the behavioral hypersensitivity (~60% responsive); mock UV-irradiated animals showed an ~27% of aversive response (Figure 3C). Similar to mechanical allodynia, the behavioral response at 8 h, 16 h, and 24 h of UV-treated animals (in the range of 36%–42%) was statistically indistinguishable from the non-treated larvae (in the range of 20%–26%). Larvae at the late third instar stage did show a slight decrease of the baseline behavioral response when compared with the middle third instar stage. We hypothesize this could be either by the increased size of the larvae (Figure 2A) or the increased thickness of the cuticle covering the body. This fact could explain why at a later stage of development the UV treatment does not induce greater mechanical sensitization, as observed 4 h post UV treatment.
Taken together, our results indicate that Drosophila larvae develop both mechanical allodynia and mechanical hyperalgesia following UV-induced tissue damage. The peak time of mechanical allodynia and hyperalgesia is the same, 4 h after UV treatment; however, mechanical hyperalgesia has a more pronounced temporal tail as it returns to baseline more slowly compared to mechanical allodynia.

Figure 1: Development of a Von Frey-like tool to evaluate mechanical nociception in Drosophila larvae. (A) Picture of a mechanical probe used to study mechanical nociception in Drosophila larvae. (B) Nitinol filaments and their relative diameters are shown to relative scale. (C) Picture of the diagonal wire cutter used to cut the nitinol filaments. (D) Smoothing the sharp edges of the cut nitinol filament with a sharpening stone. (E) Hypodermic needle used to make a hole into the wooden popsicle stick handle of the probe. The tip of the needle needs to reach at least half the height of the handle stick for secure filament insertion. (F–G) Attachment of the nitinol filament by gluing into a wooden popsicle stick handle with insertion hole. (H–L) Calibration of mechanical probes by pressing them against a scale. (M) Values of force (in mN) and pressure (in kPa) generated by different mechanical probes. The length of each nitinol filament used to construct the probes (P1–P10; P: probe) is detailed in centimeters (cm). (N) A picture of a complete set of mechanical probes, ranging from 174 kPa to 5,116 kPa. Please click here to view a larger version of this figure.
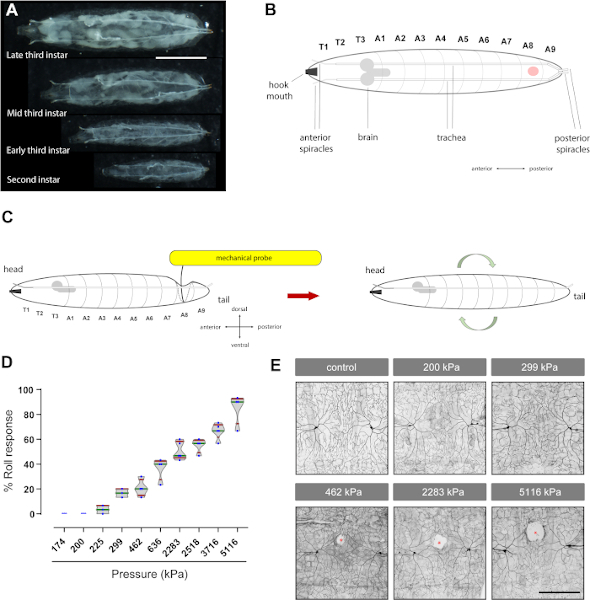
Figure 2: Mechanical nociception assay: Von Frey-like filaments generate a dose-response curve of aversive rolling behavior and cause tissue damage to sensory neurons. (A) Pictures of the different stages (second and third instar) of Drosophila larvae. Scale bar: 2 mm. (B) Cartoon of the dorsal view of the third instar Drosophila larvae. The red dot indicates the abdominal segment where the mechanical probe is applied. T: thoracic segment; A: abdominal segment. Other anatomical landmarks are labeled. (C) Cartoon of the assay: A mechanical probe is applied to the dorsal side of the larva until it bends against the surface below and is then held for 2 s. If the pressure is sufficiently high, this elicits an aversive rolling response upon release. (D) Behavioral dose response; each blue dot represents the percent of larvae that responded, with aversive rolling, to the mechanical stimulation within a set of 10 animals. Violin plot of the percent of aversive rolling behavior induced by different mechanical probes. kPa: kilopascals. Box plots represent median (green), whiskers (red) represent the 10th and 90th percentiles. (E) Tissue damage: Third instar larvae (of genotype ppk-Gal4>UAS-mCD8-GFP to label nociceptive sensory neurons) were probed at dorsal segment A8 with the indicated pressures. Fluorescently labeled paired ddaC class IV sensory neurons (across the dorsal midline) were then examined (see sections 4 and 5). White areas (red asterisks) represent gaps or tissue damage. Scale bar: 100 μm. In panel B, the larva is shown in the dorsal view, while in C it is the lateral view. Mechanical probes pressed against the dorsal cuticle-epidermis side of the larva produce a depression like-pocket at the point of contact of the tip of the probe and the surrounding areas. The solid black line curved toward the ventral side is the top of the pocket, while the dashed gray lateral line represents the lateral side and the bottom of the pocket. Please click here to view a larger version of this figure.
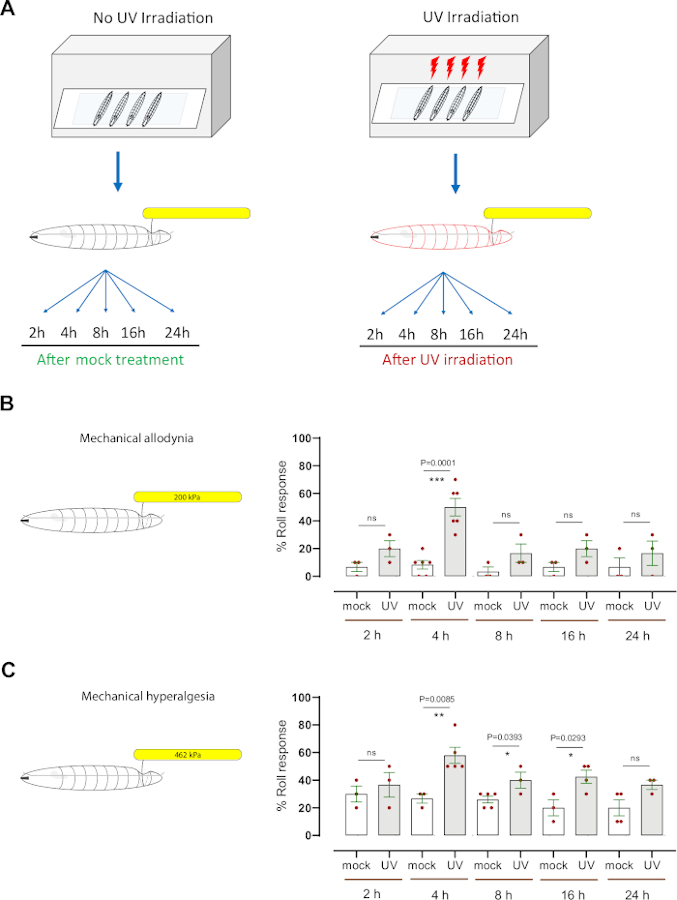
Figure 3: Mechanical hypersensitivity after UV damage. (A) Schematic of the experimental design to test sensitization. Mid third instar were mock treated (non-UV) or UV irradiated. The mechanical nociception assay was then performed at different time points (2 h, 4 h, 8 h, 16 h, and 24 h) following mock treatment or irradiation. (B) Mechanical allodynia: The percentage of larvae exhibiting aversive rolling after probing with a normally subthreshold or non-noxious mechanical stimulus (200 kPa, 1.57 mN) at the indicated time points after mock-treatment or UV irradiation. (C) Mechanical hyperalgesia: The percentage of larvae exhibiting aversive rolling after probing with a normally suprathreshold or noxious mechanical stimulus (462 kPa, 3.63 mN) at the indicated time points after mock-treatment or UV irradiation. Error bars indicate mean +/- SEM. Two-tailed unpaired t-test was used for statistical analysis: *p < 0.05, **p < 0.01; ns: not significant. Each red dot, in panels B and C, represents the mean proportion of 10 larvae, n = 3–6 sets per timepoint/condition. Please click here to view a larger version of this figure.
Video 1: Normal locomotion of Drosophila larvae. Please click here to download this video.
Video 2: Noxious mechanical stimulation of Drosophila larvae. Please click here to download this video.
Video 3: Subthreshold mechanical stimulation of Drosophila larvae. Please click here to download this video.
