This protocol will yield live pancreatic tissue slices suitable for both functionality studies and immune cell recordings. Slice appearance in both brightfield and under reflected light are shown in Figure 1A,B. As discussed, islets can be found in slices using reflected light due to their increased granularity that occurs because of their insulin content (Figure 1C) and are clearly observed compared to the background tissue when reflected light is used. Viability should be assessed following slice generation, and islets should not be recorded if more than 20% of the islet is not viable. An islet with high viability is shown in Figure 1D, whereas an example of a poorly processed slice is shown in Supplemental Figure 1. Islets with low viability will have heavy SYTOX Blue staining, and the tissue will be covered with the stained nuclei of dead cells. Additionally, calcein-AM and Ca2+ indicators such as the Oregon Green 488 BAPTA-1, AM used here and Fluo-4-AM will not load well in dead cells. Islets should be selected for Ca2+ recordings if they are viable and if the indicator is loaded throughout the islet. Ca2+ indicator loading is indicative of cell viability as both Ca2+ indicators discussed in this protocol (the Oregon Green 488 BAPTA-1, AM and Fluo-4-AM) are loaded in cells through the same mechanism as the viability dye, calcein-AM.
For both the Ca2+ indicator dyes and calcein-AM, when the stains are loaded into cells, the acetoxymethyl ester is hydrolyzed within the cell, and the molecule becomes membrane impermeable19. Another positive indicator for viability is observable basal activity throughout the islet with cells flashing on and off. Basal activity should also be observable in the exocrine tissue to a lesser degree. Although mouse tissue tends to have less visible basal activity than human tissue, it is still present. An islet from a slice made from a NOD.Rag1-/- mouse pancreas is shown in Figure 2A. As mentioned above, the Oregon Green 488 BAPTA-1, AM used here has a lower fluorescence intensity increase upon binding Ca2+ (~14-fold) than Fluo-4 (~100-fold). However, the Oregon Green 488 BAPTA-1, AM has the advantage of a lower calcium dissociation constant (Kd = 170 nM) than Fluo-4 (Kd = 335 nM), resulting in the Oregon Green 488 BAPTA-1, AM being more sensitive to lower concentrations of cytosolic Ca2+. However, responses are still quantifiable, as shown by Figure 2B. Examples of an islet within a control human pancreatic tissue slice at rest and of one exhibiting a strong high glucose response are shown in Figure 2C and Supplemental Video 1. Fluo-4-AM dye is loaded well and is visible throughout the islet at low glucose concentrations. As discussed above, a typical occurrence is for a percentage of cells to load large amounts of dye and appear very bright. Moreover, the image parameters have been set for this recording so that most of the cells within the islet do not appear too bright at low glucose concentrations. This enables the detector to pick up on the increases in brightness that occur during changes in intracellular Ca2+ concentrations in response to high glucose levels. The quantification of the fluorescence of individual cells during this response is shown in Figure 2D, with the expected peak following the high glucose stimulation. ImageJ software was used to calculate the staining intensity of Fluo-4-AM and the Oregon Green 488 BAPTA-1, AM by manually selecting ROIs. The fold-increase in fluorescence intensity for each ROI was calculated by normalizing the fluorescence values at later timepoints using the initial fluorescence values of the cells (F/F0).
Dithizone stains the islets red and is visible under a brightfield stereomicroscope. Intact islets and islets that are beginning to fall apart because of T1D onset can both be observed using this dye (Figure 3A,B). Islets can be found using reflected light (Figure 3C) and may begin to lose granularity due to immune cell infiltration and cell death (Figure 3D). Multiple CD3-positive cells can be seen infiltrating the islet in Figure 3D. Immune cell populations can be identified more specifically using CD8 antibody and insulin-tetramer staining. Imaging can then be applied to identify cells that co-stain for both markers (Figure 3E). The co-staining of the immune cells infiltrating the islet in Figure 3D indicates that the cells are effector T cells that are specifically targeting the insulin antigen. The CD8 co-stain is essential to distinguish that the areas that stain positive for tetramer are immune cells. The tetramer should not be used alone without an immune cell co-stain. A staining comparison of the mouse CD8 antibody and the isotype control Rat IgG2a, κ can be found in Supplemental Figure 2. An additional comparison of a control tetramer for lymphocytic choriomeningitis virus (LCMV) tetramer and the insulin tetramer can be found in Supplemental Figure 3. Some T cells remain stationary throughout the recording, many move slightly within a small area of the islet, and others are very mobile and can be seen moving throughout the islet and exocrine tissue. It is not unusual to see T cells exhibiting multiple mobility types within the same recording.
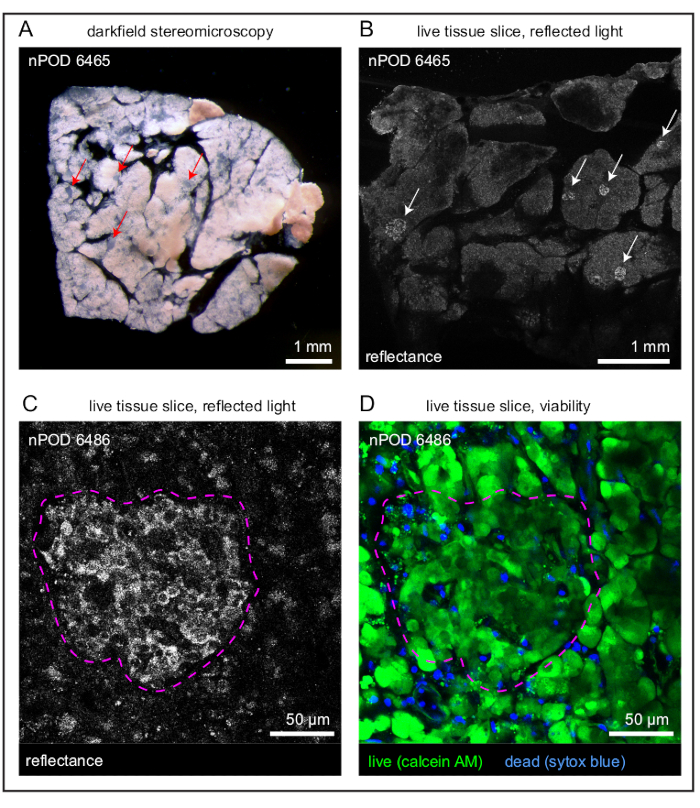
Figure 1: Overview of slices and individual islets. (A) Darkfield stereomicroscopy image of a live human pancreatic tissue slice with islets indicated by red arrows. (B) Reflected light image of a live human pancreatic tissue slice with islets indicated by white arrows. (C) Reflected light image of an islet (outlined in magenta) within a live human pancreatic tissue slice. (D) Viability staining of a high-viability islet (outlined in magenta) within a live human pancreatic tissue slice. Live cells are indicated in green and dead cells in blue. Scale bars (A, B) = 1 mm; scale bars (C, D) = 50 µm. Abbreviation: AM = acetoxymethyl ester. Please click here to view a larger version of this figure.
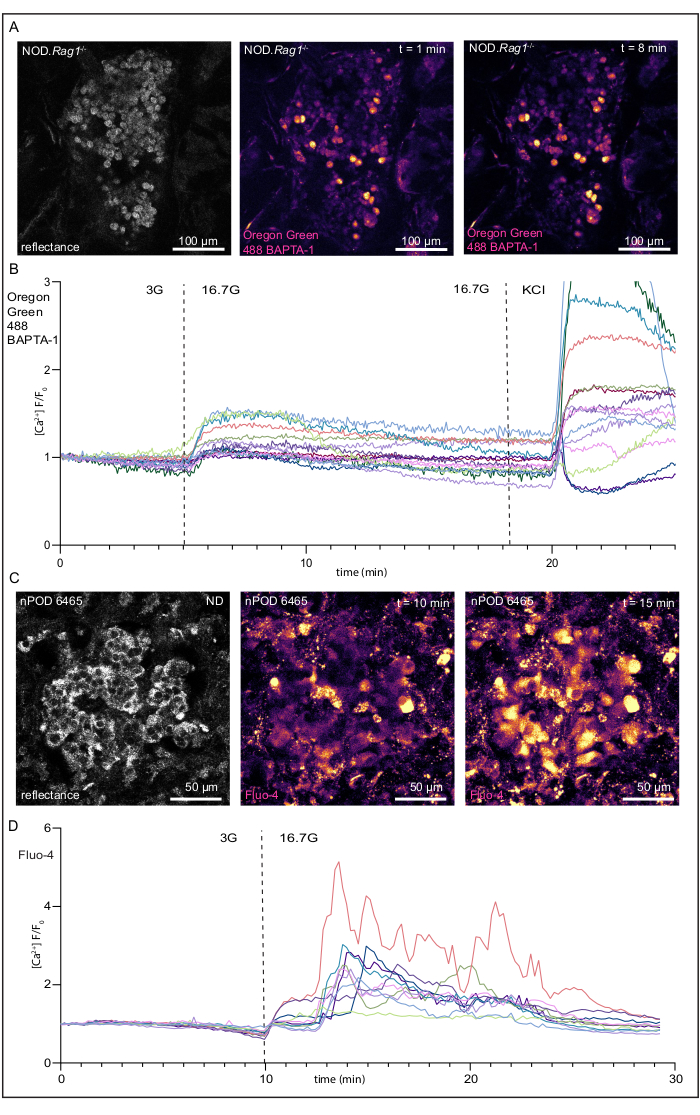
Figure 2: Recordings of changes in intracellular Ca2+ concentrations and responses to high glucose concentration of a live NOD.Rag1-/- mouse pancreatic tissue slice and human pancreatic tissue slice from a donor without diabetes. (A) Images of an islet within a live NOD.Rag1-/- mouse pancreatic slice loaded with a Oregon Green 488 BAPTA-1, AM (see the Table of Materials) undergoing glucose stimulation. From left to right, a reflected light image of the islet, the islet in low glucose, and the islet in high glucose. (B) Fluorescence traces of the Ca2+ response of an islet within a live NOD.Rag1-/- tissue slice with the expected response to high glucose concentration [KRBH with 16.7 mM D-glucose (16.7G)] and KCl [KRBH with 30 mM KCl and 3 mM D-glucose]. (C) Images of an islet within a live human pancreatic slice loaded with Fluo-4-AM undergoing glucose stimulation. From left to right, a reflected light image of the islet, the islet in low glucose, and the islet in high glucose. (D) Fluorescence traces of the Ca2+ response of an islet within a live human pancreas tissue slice with the expected response to KRBH with 16.7 mM D-glucose (16.7G). Scale bars (A) = 100 µm; scale bars (C) = 50 µm. Abbreviations: KRBH = Krebs-Ringer bicarbonate buffer; KCl = potassium chloride; NOD.Rag1-/- = non-obese diabetic-recombination activating gene-1-null; NOD.Rag1-/-.AI4α/β = T cell receptor transgenic (AI4) mouse strain. Please click here to view a larger version of this figure.
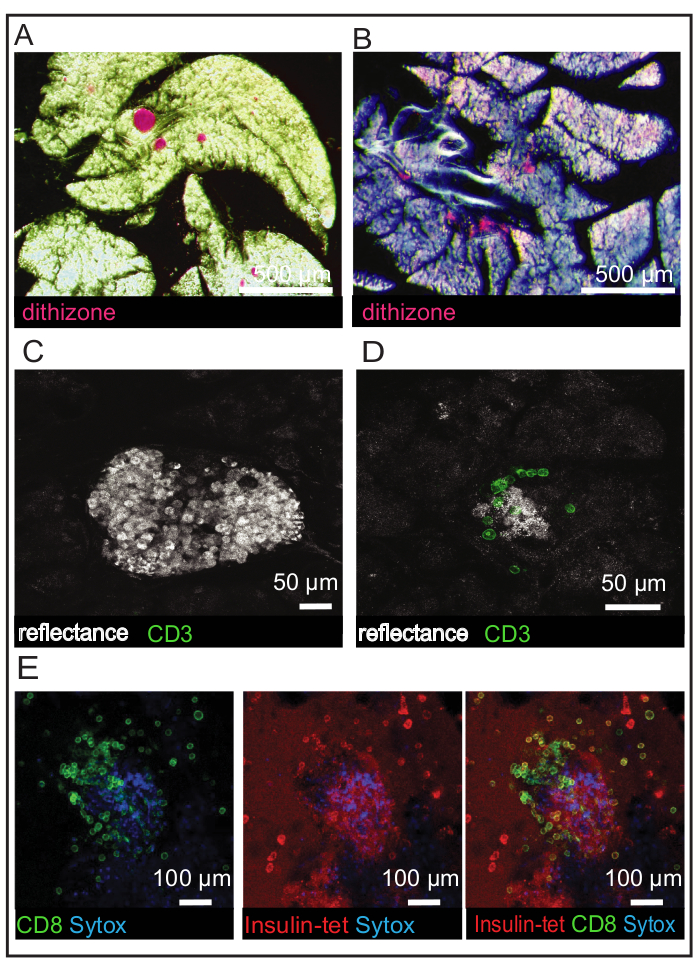
Figure 3: Identification of islets and immune cell populations in NOD.Rag1-/- and NOD.Rag1-/-.AI4α/β mouse slices. (A) Dithizone staining of islets in a NOD.Rag1-/- mouse slice with the islets indicated in red. (B) Dithizone staining of islets in a NOD.Rag1-/-.AI4α/β mouse slice with the islets indicated in red. Islets are losing their shape due to disease onset. (C) Reflected light image of an islet in a NOD.Rag1-/- mouse slice. (D) Reflected light image of an islet in a NOD.Rag1-/-.AI4α/β mouse slice with CD3 antibody staining (green). (E) Viability staining of dead cells (blue) and immune cell staining (CD8 in green and insulin tetramer in red) in a NOD.Rag1-/-.AI4α/β mouse slice. Scale bars (A) = 500 µm; scale bars (B) = 50 µm; scale bars (C) = 100 µm. Abbreviations: NOD.Rag1-/- = non-obese diabetic-recombination activating gene-1-null; NOD.Rag1-/-.AI4α/β = T cell receptor transgenic (AI4) mouse strain; CD = cluster of differentiation; insulin-tet = insulin tetramer. Please click here to view a larger version of this figure.
Supplemental Figure 1: NOD.Rag1-/- mouse pancreatic slice following improper preparation without trypsin inhibitor and an overnight incubation at 37 °C. (A) Darkfield stereomicroscopy image of a live NOD.Rag1-/- mouse pancreatic tissue slice; scale bar = 1 mm. (B) Reflected light image of a live mouse pancreatic tissue slice; scale bar = 50 µm. (C) Viability staining of low-viability tissue. Dead cells are indicated in blue; scale bar = 50 µm. Abbreviation: NOD.Rag1-/- = non-obese diabetic-recombination activating gene-1-null. Please click here to download this File.
Supplemental Figure 2: Rat IgG2a, κ isotype control antibody (left) and rat anti-mouse CD8 antibody (right) staining comparison in NOD.Rag1-/-.AI4α/β mouse slices. (A) Reflected light images of live NOD.Rag1-/-.AI4α/β mouse pancreatic tissue slices showing an islet (left) and blood vessel (right). (B) Antibody staining of live NOD.Rag1-/-.AI4α/β mouse pancreatic tissue slices. (C) Overlay of the reflected light and antibody channels. Scale bars for control antibody (left panels) = 20 µm; scale bars for CD8 antibody (right panels) = 50 µm. Abbreviations: NOD.Rag1-/- = non-obese diabetic-recombination activating gene-1-null; NOD.Rag1-/-.AI4α/β = T cell receptor transgenic (AI4) mouse strain; CD = cluster of differentiation; IgG = immunoglobulin G. Please click here to download this File.
Supplemental Figure 3: Lymphocytic choriomeningitis virus tetramer (left) and insulin tetramer (right) staining comparison in NOD.Rag1-/-.AI4α/β mouse slices. (A) Reflected light images of live NOD.Rag1-/-.AI4α/β mouse pancreas tissue slices showing a blood vessel in exocrine tissue (left) and islets (right). (B) Tetramer staining of a live NOD.Rag1-/-.AI4α/β mouse tissue slices. (C) Overlay of the reflected light and tetramer channels. Abbreviations: NOD.Rag1-/- = non-obese diabetic-recombination activating gene-1-null; NOD.Rag1-/-.AI4α/β = T cell receptor transgenic (AI4) mouse strain; LCMV = lymphocytic choriomeningitis virus; insulin-tet = insulin tetramer. Please click here to download this File.
Supplemental Video 1: Recording of cytosolic Ca2+ detected with Fluo-4 in response to high glucose stimulation in a human pancreatic tissue slice from a control donor without diabetes. Cells within the tissue can be observed to exhibit basal Fluo-4 activity in a low glucose solution (3.0 mM), followed by an increase in Fluo-4 fluorescence intensity in response to a stimulation with high glucose (16.7 mM). The video corresponds to the still images and traces shown in Figure 2C,D. Please click here to download this Video.
Supplemental Table 1: Please click here to download this Table.
