To induce AAV-mediated overexpression of a tumor-restraining stromal gene, Islr4,25,43,44, in hepatocytes, we intravenously injected Islr-encoding AAV8. 1.0 x 1011 viral genomes (vg) of AAV8-Islr, or as a control, AAV8-mRuby2, was injected into the adult mouse tail vein (Figure 1A). Two weeks after the tail vein injection, livers were harvested to validate the overexpression of Islr in hepatocytes. We performed RNAscope in situ hybridization45 and confirmed that 3,3'-Diaminobenzidine (DAB)+ signals were observed throughout the liver (Figure 1B). No DAB+ signals were detected in the liver from an AAV8-mRuby2-treated mouse.
We practiced the portal vein injection procedure by injecting India ink (1:1000 dilution in PBS). After making an incision into the upper abdomen, the intestines were gently taken out from the abdominal space to allow visualization of the portal vein (Figure 2A). Portal vein injection of India ink delivered the ink throughout the liver, but not to the lung (Figure 2B). If the ink is mistakenly injected into other vessels such as the inferior vena cava (IVC) or abdominal aorta, systemic circulation of the ink alters the lung color to black. India ink also helps identify the amount of leakage from the portal vein, which indicates pancreatic or peritoneal dissemination.
Two weeks after the tail vein injection of AAV-mRuby2, ApcΔ/Δ and Trp53Δ/Δ colon cancer organoids (AP tumoroids) were dissociated to single cells and injected into the mouse portal vein (Figure 3A). To confirm metastatic tumor growth and evaluate histology, we collected the livers 3-4 weeks after the portal vein injection (i.e., at a timed point rather than at a humane endpoint), before prominent necrosis confounds histopathological analyses. Macroscopically, the portal vein injection of tumoroids resulted in multiple white tumor nodules in the liver (Figure 3B). We found that intra-splenic injection of the same cell number of tumoroids did not generate a large metastatic tumor mass. This suggests that the portal vein injection approach more efficiently induced liver metastases compared to the intra-splenic injection model. Hematoxylin and eosin staining of the CRC hepatic metastases induced by the portal vein injection demonstrated histopathology of moderately differentiated tubular adenocarcinoma accompanied with a desmoplastic stromal reaction and necrosis (Figure 3C). This stroma-rich histology faithfully recapitulated that of human CRC liver metastases (Figure 3C), making this model suitable for translational research investigating the metastatic tumor stroma. Moreover, immunohistochemistry for alpha-smooth muscle actin (αSMA), a well-established marker for CAFs, showed that about 7% of tumor areas were αSMA-positive, confirming the presence of CAFs in this mouse model (Figure 3D,E). Picro-Sirius red staining that stains collagen46 demonstrated abundant ECM in the tumor mesenchyme with approximately 13% of tumor areas positive for collagen (Figure 3D,E). Immunohistochemistry for EPCAM, an epithelial lineage marker, revealed that the metastatic CRC showed tumor budding (a single tumor cell or a cell cluster of up to four tumor cells) that is a characteristic of poor prognosis colorectal cancer47 (Figure 3F). Ki67 labeling index in the metastatic CRC was approximately 80%, indicating that most tumor cells are mitotically active (Figure 3G,H).
We performed mouse survival and tumor growth kinetics analyses in this preclinical model. The mice in our first pilot cohort showed a median survival of 57 days after tumoroid injection into the portal vein (N = 4 mice; Figure 4A). This was later replicated in larger control AAV8-mRuby2-injected groups25. At humane endpoint (as shown in NOTE, step 5.2), 3 out of 4 mice in the pilot group demonstrated ascites, which is also observed in patients with advanced liver metastasis48. To assess the tumor growth, in vivo imaging system (IVIS) was used to measure tumoroid-derived luminescence (Figure 4B,C). The bioluminescence signals were observed within the upper abdomen, suggesting liver-specific tumor growth (Figure 4B). If IVIS signals are observed in the lower abdomen, this indicates peritoneal dissemination or secondary metastases to abdominal organs. The weekly in vivo imaging allowed for a longitudinal assessment of tumor growth in each mouse (Figure 4C), making it easier to monitor a therapeutic effect in our subsequent larger study. The tumor take rate was 100% (4/4 mice) as assessed by IVIS signals. In this experiment, no macroscopically apparent lung metastasis was observed at the time of tissue collection (0/4 mice).
Finally, we investigated whether AAV-mediated hepatocyte-directed delivery of a cancer-restraining CAF gene, Islr4,25, could inhibit CRC hepatic metastasis growth in this preclinical mouse model. Notably, AAV8-Islr-treated mice showed improved mouse survival and decreased IVIS signals from tumors (Figure 5A–C)25. Immunohistochemistry for phosphorylated Smad1/5/8 demonstrated that hepatocyte overexpression of ISLR, a BMP signaling potentiator, augmented BMP signaling in metastatic tumors (Figure 5D). Treatment with AAV8-Islr decreased the number of Ki67+ proliferating cells in the CRC hepatic metastasis (Figure 5E). For full description of the results, refer to Kobayashi et al., Gastroenterology, 202125. Our collective data indicate that AAV8-mediated delivery of a tumor-inhibitory gene to hepatocytes, a vital constituent of the liver metastatic tumor stroma29, could be an effective preventive/therapeutic approach for CRC liver metastases (Figure 5F).
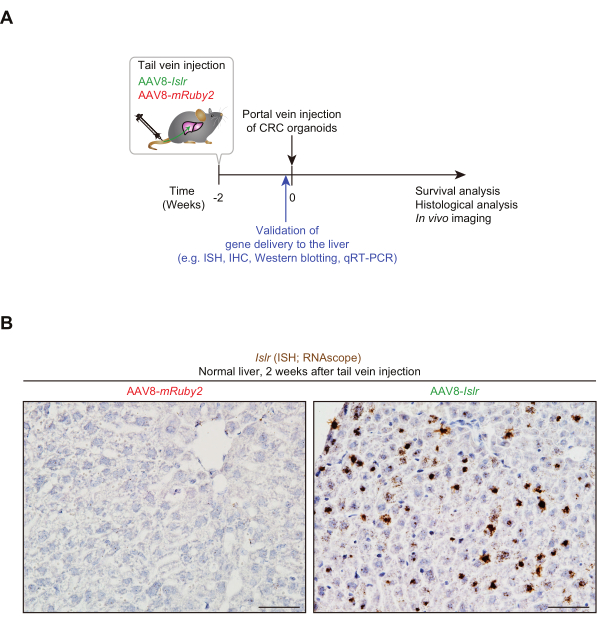
Figure 1: Tail vein-administered AAV8-Islr generates Islr overexpression in the liver. (A) Experimental scheme showing the tail vein injection of AAV8, followed by portal vein injection of colorectal cancer (CRC) organoids. ISH, in situ hybridization; IHC, immunohistochemistry; qRT-PCR, quantitative real-time polymerase chain reaction. (B) Representative pictures. RNA in situ hybridization for Islr was performed using livers from AAV8-Islr or AAV8-mRuby2-treated mice. The livers were collected 2 weeks after tail vein injection. Scale bars, 50 µm. Please click here to view a larger version of this figure.
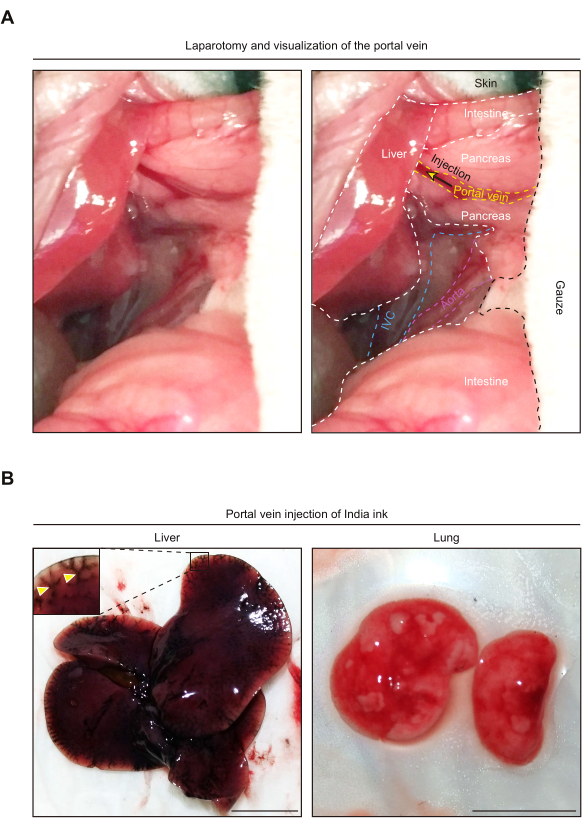
Figure 2: Portal vein injection of India ink results in its delivery to the liver, but not the lung. (A) Representative pictures showing anatomy of the upper abdomen after laparotomy. Note that, for visualization of the portal vein, the intestines are taken outside the abdominal cavity. The right-hand side picture shows an anatomical annotation of each organ and vessel. The yellow arrow denotes the injection site. IVC, Inferior vena cava. (B) Representative pictures of the liver and lung following portal vein injection of India ink. The yellow arrowheads indicate vessels stained with the India ink. Scale bars, 1 cm. Please click here to view a larger version of this figure.
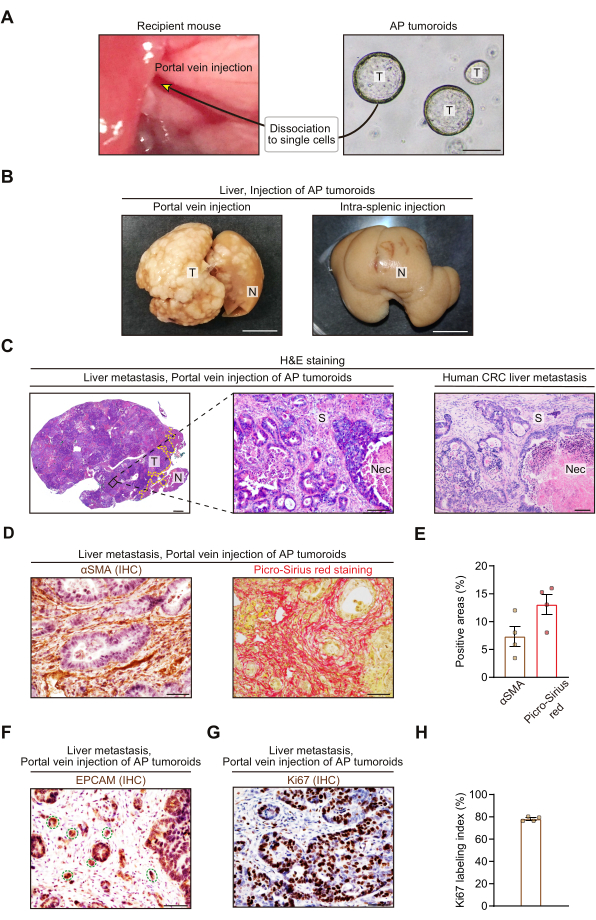
Figure 3: Portal vein injection of colorectal cancer organoids generates stroma-rich liver metastases. (A) Experimental scheme showing portal vein injection of ApcΔ/Δ and Trp53Δ/Δ colon cancer organoids (AP tumoroids). Scale bar, 200 µm. T, tumoroids. (B) Representative macroscopic pictures of the livers that were collected 3-4 weeks after portal vein injection (left) or intra-splenic injection (right). 5.0 x 105 single cells from AP tumoroids were injected into the portal vein or spleen. Intra-splenic injection was performed as described13. White nodules indicate tumors. T, Tumor; N, Normal liver. (C) Representative hematoxylin and eosin (H&E) staining pictures of the CRC liver metastases from the portal vein injection mouse model (left and middle) and human (right). T, Tumor; N, Normal liver; S, Stroma; Nec, Necrosis. The yellow dotted line indicates a border between the tumor and normal liver (left). (D and E) Immunohistochemistry (IHC) for alpha-smooth muscle actin (αSMA; left) and Picro-Sirius red staining (right). (D) Representative pictures. (E) Quantification of αSMA-positive areas (left) and Picro-Sirius red-positive areas (right). N = 4 mice, 5 HPFs (High Power Fields; 400x)/mouse. (F) Representative picture showing immunohistochemistry for EPCAM, an epithelial cell marker. The green dotted lines denote tumor budding. (G and H) Immunohistochemistry for Ki67, a cell proliferation marker. (G) Representative picture. (H) Percentage of Ki67+ cells in total epithelial cells. Epithelial cells were visualized by hematoxylin counterstaining. N = 4 mice, 5 HPFs/mouse. In (B)-(H), all mouse liver tissues were collected 3-4 weeks after injection of AP tumoroids. Mean ± S.E.M. Each dot represents an average value of 5 HPFs from a mouse (E and H). Scale bars represent 1 cm (B), 1 mm (C; left), 100 µm (C; middle and right), and 50 µm (D, F, and G). Note that the study with human tissues was approved by the Ethics Committee of Nagoya University Graduate School of Medicine (2017-0127). Please click here to view a larger version of this figure.
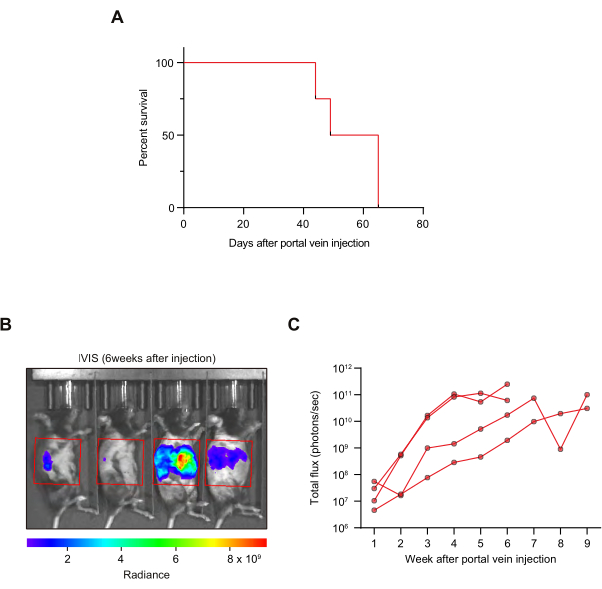
Figure 4: Survival analysis and tumor growth kinetic analysis by in vivo imaging. (A) Kaplan-Meier survival curves. N = 4 mice. (B,C) In vivo imaging system (IVIS) was used to evaluate tumor growth kinetics. (B) A representative picture. The area in the red box was used for quantification. (C) Growth kinetics. Luciferase signals from each mouse are shown. N = 4 mice. Please click here to view a larger version of this figure.
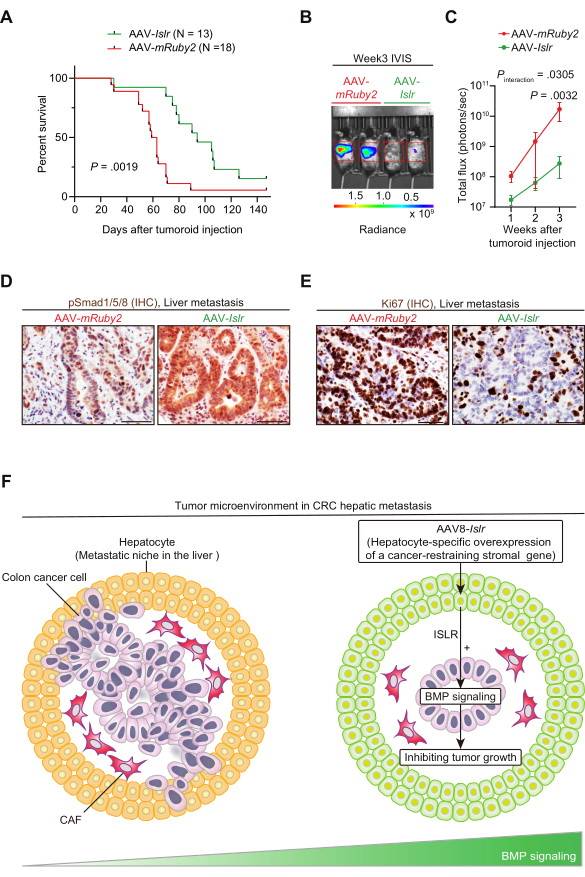
Figure 5: AAV8-mediated gene delivery of Islr to hepatocytes increases BMP signaling, reduces tumor proliferation, and improves mouse survival in a portal vein injection model of CRC liver metastasis. (A) Kaplan-Meier survival curve. Two weeks after tail vein injection of AAV-Islr or AAV-mRuby2, portal vein injection of CRC tumoroids was performed. (B and C) Tumoroid-derived luciferase signals were evaluated using IVIS. (B) Representative picture. (C) Quantification of IVIS signals. N = 5 (AAV-mRuby2) and 8 (AAV-Islr) mice. Mean ± S.E.M. (D) Representative pictures. Immunohistochemistry (IHC) for phosphorylated Smad1/5/8 (pSmad1/5/8). Smad1/5/8 is a downstream molecule of bone morphogenetic protein (BMP) signaling that is phosphorylated upon BMP signaling activation49. N = 4 mice each. (E) Representative pictures. Immunohistochemistry for Ki67. N = 4 mice each. (F) Graphical summary. AAV-mediated hepatocyte-directed gene delivery of a cancer-restraining stromal gene might serve as a therapeutic strategy to inhibit CRC metastagenesis. AAV8-mediated overexpression of Islr in hepatocytes remodeled the metastatic niche by augmenting BMP signaling and limited CRC metastasis progression. For detailed information, refer to Kobayashi et al., Gastroenterology, 202125. Log-rank test (A), and two-way repeated-measures ANOVA (analysis of variance) with post-hoc Sidak's multiple comparison test at Week 3 (C). Scale bars, 50 µm. Figure 5A–C has been reprinted from Gastroenterology, Vol 160(4), Kobayashi et al., The Balance of Stromal BMP Signaling Mediated by GREM1 and ISLR Drives Colorectal Carcinogenesis, Pages 1224-1239.e30, Copyright (2021), with permission from Elsevier. Please click here to view a larger version of this figure.
