The roller culture conditions described for E7.5 embryos (late-gastrulation stage) support constant and normal embryo growth with an average efficiency close to 75% after 4 culture days (Figure 2 and Table 1). The efficiency of embryo development may vary across diverse mouse genetic backgrounds but is consistently robust (Figure 2C). Supplementation with HBS instead of HCS yields an efficiency of ~68% after 4 days of ex utero culture, depending on the genetic background of the mice (Figure 2D and Table 2). The embryos developed ex utero recapitulate proper development until approximately the 44-somite stage. Afterward, the embryos present embryonic abnormalities due to the absence of the allantoic placenta, resulting in insufficient oxygenation and nutrient supply given the increased body size at this stage.
Development of E6.5 embryos (early-streak) in static plates is correctly recapitulated with an efficiency of >90% until the early somite-stage E8.5, using EUCM with both HCS and HBS (Figure 3, Table 3, and Table 4) (see8,13 for a detailed description of embryo staging between E5.5 to E8.5). Ex utero culture from gastrulation to advanced organogenesis by combining cultures on static plates followed by the roller culture in a constant 21% oxygen atmosphere gives an estimated efficiency of proper development of 55% and 26% to the 44-somite stage, using HCS and HBS, respectively (Figure 3A, Table 3, and Table 4). There is a delay of 1-2 somite pairs in these embryos compared to embryos developed in utero. The greatest drop in efficiency occurs at the transition from E8.5 to E9.5 due to failure of axial turning and closure of the neural tube.
Cultures starting from E5.5 pregastrulating embryos show efficiency of proper development to the early-somite stage (E8.5) of approximately 46%, and nearly 17% of the embryos will complete proper development after six days of culture after being transferred to the roller culture (Figure 4 and Table 5). Extended ex utero culture prolongs the developmental delay in the embryos, with embryos explanted at E5.5 showing a delay of 2-4 pairs of somites compared to in vivo embryos. Nevertheless, morphogenesis and tissue development proceed properly until approximately the 42-somite stage.
The most common defects seen in the embryos for cultures initiated from E7.5, E6.5, and E5.5 are exemplified in Figure 5A–C. At the time of dissection, embryos with even minor damage to the epiblast or the extraembryonic region, as well as embryos retaining the Reichert's membrane, should be discarded. Likewise, early embryos will not grow properly (see Figure 5B for dead embryos) or display severe developmental delays (see Figure 5B for a delayed embryo). Attachment of the embryonic epiblast to the surface of the plate will affect development depending on the position and grade of attachment. Attachment of a part of the epiblast or the whole embryo will cause the failure of further development (see Figure 5B for an attached embryo).
The main abnormalities observed in the percentage of defective embryos at E8.5 (early somite stage) are the development of the posterior region outside the yolk sac or defects in the growth of the neural folds (Figure 5A,B). In the case of cultures started at E5.5, a frequently observed developmental defect is the presence of a small, underdeveloped epiblast (Figure 5C). At the time equivalent to E9.5, defects in the closure of the neural folds, failure of axial turning, or a deficiency in brain growth represent the most commonly observed abnormalities (Figure 5). The most frequently observed developmental defects at E10.5/E11 are anomalies in the head region, disruption of normal blood circulation in the yolk sac, and pericardial effusion (Figure 5). Rupture of one main blood vessel and blood outflow may cause subsequent death of the embryo. Notably, proper growth of the embryo itself might be reached even in the absence of evident yolk sac circulation. Embryos kept in culture beyond the stage equivalent to E11 exhibit body shrinkage and death after few hours due to a lack of proper tissue oxygenation.

Figure 1: Gas and pressure regulation system adapted to a roller culture incubator. (A) Top view of the gas regulation module connected to the roller culture incubator. N2 and CO2 enter the gas regulator to allow precise control of the oxygen/CO2 concentrations and gas pressure. The gases are directed towards the mixing box, in which they are mixed by a centrifugal blower and injected into the incubator by a pump that generates positive pressure. The gas flows through the inlet into a water bottle and later to the sealed bottles. (B) Internal configuration of the electronic module for gas and pressure regulation. The voltage value set on the pressure transmitter regulates the pressure generated by the pump inside the gas mixing box (5-6 V to attain pressure of 6-7 psi in this specific model). Please click here to view a larger version of this figure.
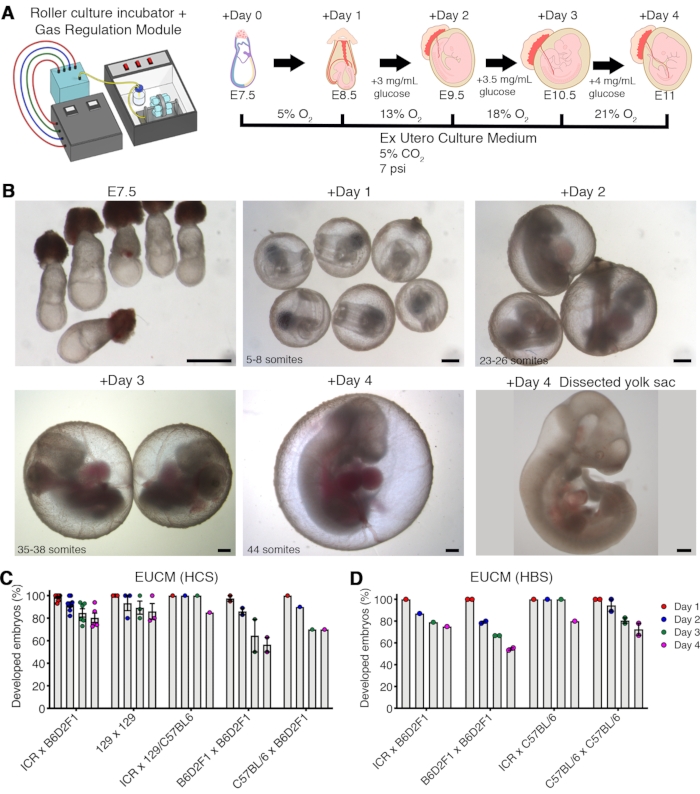
Figure 2: Ex utero culture platform supports growth of E7.5 embryos until advanced organogenesis. (A) Diagram depicting the E7.5 ex utero embryo culture protocol. (B) Representative bright-field images of groups of cultured embryos developing ex utero over 4 days, from late gastrulation (E7.5) to the 44-somite stage (E11). The typical variation in somite number assessed every 24 h is indicated. Scale bars = 500 µm. (C, D) Percentage of normally developed embryos at 1-4 days of culture starting from E7.5 divided by mouse parental strains and serum supplementation (C, human umbilical cord blood serum; D, human adult blood serum). Panel A has been modified from 13. Abbreviations: EUCM = ex utero embryo culture medium; HCS = human umbilical cord blood serum; HBS = human adult blood serum. Please click here to view a larger version of this figure.
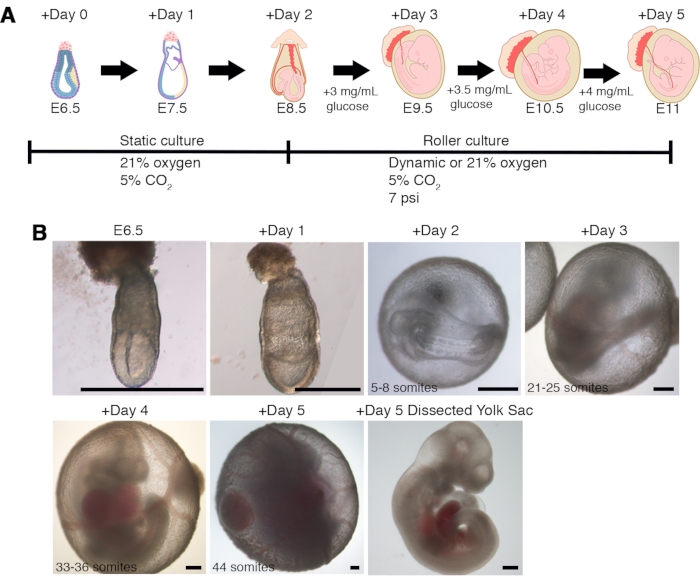
Figure 3. Extended ex utero culture protocol for growing E6.5 early-gastrulating mouse embryos until late organogenesis. (A) Schematic illustration of the extended ex utero culture protocol combining static plates and rotating bottles systems. (B) Bright-field images of embryos cultured ex utero for five days from E6.5 to the 44-somite stage. The typical variation in somite number assessed every 24 h is indicated. Scale bars = 500 µm. Please click here to view a larger version of this figure.
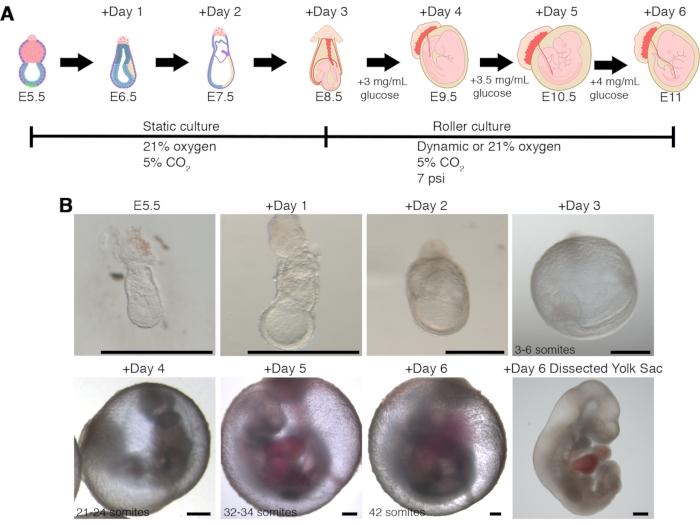
Figure 4: Continuous ex utero culture of pregastrulation mouse embryos from E5.5 until late organogenesis stages. (A) Schematic depiction of the ex utero culture the protocol for E5.5 embryos.(B) Representative bright-field images of embryos cultured ex utero for six days from E5.5 until the 42-somite stage. The typical variation in somite number assessed every 24 h is indicated. Scale bars = 500 µm. Please click here to view a larger version of this figure.
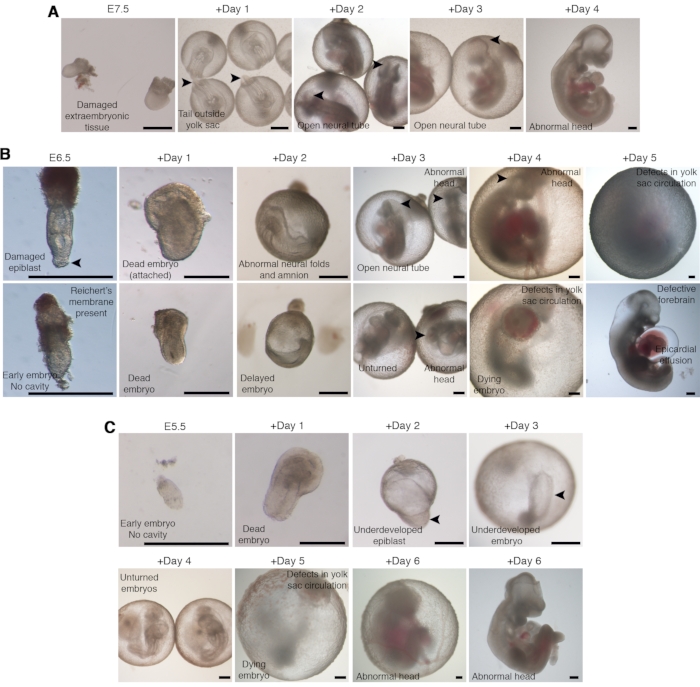
Figure 5: Representative developmental defects observed in embryos cultured ex utero. (A–C) Bright-field microscopy images of abnormal mouse embryos grown ex utero starting from E7.5 (A), E6.5 (B), or E5.5 (C). A general description of the defect is provided on each image. Scale bars = 500 µm. Please click here to view a larger version of this figure.
Table 1: Efficiency of proper development of embryos isolated at E7.5 days post coitum. The embryos were cultured ex utero for 4 days in EUCM (25% Human Umbilical Cord Blood Serum). [-] indicates cultures not continued due to experimental requirements. Please click here to download this Table.
Table 2: Efficiency of proper development of embryos isolated at E7.5, cultured ex utero for 4 days, replacing Human Umbilical Cord Blood Serum with freshly isolated Human Adult Blood Serum. [-] indicates cultures not continued due to experimental requirements. Please click here to download this Table.
Table 3: Efficiency of proper development of embryos (B6D2F1/ICR) isolated at E6.5 and cultured ex utero for 5 days using EUCM (25% Human Umbilical Cord Blood Serum). The ex utero culture was done in static culture for two days (21% O2) followed by three days in rotating bottles at 21% O2. [-] indicates cultures not continued due to experimental requirements. Abbreviation: NA = not acquired. Please click here to download this Table.
Table 4: Efficiency of proper development of embryos (B6D2F1/ICR) isolated at E6.5 and cultured ex utero for 5 days using EUCM (replacing Human Umbilical Cord Blood Serum with freshly isolated Human Adult Blood Serum). The embryos were developed in static culture for two days (21% O2) followed by three days in rotating bottles at 21% O2. [-] indicates cultures not continued due to experimental requirements. Please click here to download this Table.
Table 5: Efficiency of proper development of embryos (B6D2F1/ICR) isolated at E5.5 and cultured ex utero for 6 days using EUCM (25% Human Umbilical Cord Blood Serum). The embryos were developed in static culture for three days (21% O2) followed by three days in rotating bottles at 21% O2. [-] indicates cultures not continued due to experimental requirements. Please click here to download this Table.
