This protocol enables the derivation of human lung organoids with a high success rate. Fresh human lung tissue is minced into small pieces, and then decomposed with collagenase. The resultant single cells are embedded in the basement matrix and incubated in the lung organoid expansion medium supplemented with a cocktail of niche factors for the outgrowth of epithelial stem cells (step 1.1.2). Figure 1 shows the microphotograph of freshly isolated lung cells embedded in reduced growth factor basement membrane matrix Type 2 (BME; Figure 1A, left). Cystic organoids appear and enlarge over time (Figure 1A, right). Meanwhile, the unrelated cells undergo cell death gradually. Fibroblasts are present in the culture during the first or second passages. Afterward, the culture contains epithelial organoids exclusively, which are lung organoids derived from epithelial stem cells present in the primary lung tissues. These lung organoids are passaged every 2-3 weeks by mechanical shearing at a ratio of 1:3 to 1:5 or by trypsinization at a ratio of 1:5 to 1:10 (steps 2.2-2.3). The representative microphotographs of lung organoids after the fourth passage are shown in Figure 1B. After mechanical shearing, the organoid fragments embedded in BME form cystic domains within a couple of hours (Figure 1B, left). A microphotograph of the same field on day 5 (Figure 1B, right) shows organoids growing over time. These expanding human lung organoids harbor all the four major airway epithelial cell types, including ACCTUB+ or FOXJ1+ ciliated cell, P63+ basal cell, CC10+ club cell, and MUC5AC+ goblet cell18 (Figure 1C), in a premature state. Notably, these human lung organoids can be consecutively and stably passaged for over 1 year. When maintained within the basement matrix, lung organoids are most likely to show an apical-in polarity, less than 2%-3% of lung organoids show an apical-out polarity13. As a result, cell apexes are not readily accessible unless the 3D organoids are sheared open.
However, compared to the native human airway epithelium, these long-term expandable lung organoids are not sufficiently mature since the dominant cell population in the native human airway epithelium, ciliated cell, is under-represented in the lung organoid. We then defined a proximal differentiation (PD) medium to improve the maturation status of human lung organoids. The organoids incubated in the expansion medium and the PD medium developed distinct morphology over time (Figure 2A). Motile cilia were more abundant in the organoids in the PD medium than those in the expansion medium. After 2 weeks of differentiation culture in the PD medium, motile cilia are discernible in every single organoid (Figure 2B and Supplementary Video 1). Interestingly, the beating cilia drive the cell debris and excreted mucin inside the organoid lumen to swirl unidirectionally, which adequately recapitulates the mucociliary escalator to remove the inhaled particles (Supplementary Video 1), an important self-clearing mechanism of the human airways. We demonstrate that the ciliated cells increased dramatically to around 50% in the differentiated organoids as compared to the original lung organoids. To assess the percentages of four types of epithelial cells, the 2D airway organoids were analyzed by flow cytometry. Briefly, the organoids were dissociated with 10 mM EDTA for 60 min at 37 °C, fixed with 4% PFA, and permeabilized with 0.1% surfactant. Subsequently, the cells were incubated with primary antibodies (see Table of Materials) for 1 h at 4 °C followed by staining with secondary antibodies. A FACS system was used to analyze the samples. The flow cytometry analysis demonstrated differentiated organoids accommodate four airway epithelial cell types (Figure 2C). Therefore, we developed a proximal differentiation protocol to generate airway organoids that can faithfully simulate the human airway epithelium to a near-physiological level.
To enable the organoid apical surface to be readily accessible and better model the human airway epithelium exposure to respiratory pathogens, we generated 2D monolayers of airway organoids. After 2 weeks of differentiation culture, 2D airway organoids developed an intact epithelial barrier (Figure 3A,B). We also performed a dextran blockage assay to assess the integrity of epithelial barrier formed in 2D airway organoids. On day 10 after culture in transwell inserts, fluorescein isothiocyanate-dextran (MW 10,000) was added in the medium of the top chamber and incubated at 37 °C for 4 h. The media in the top and bottom chambers were harvested for a fluorescence assay. Dextran blockage index refers to the fluorescence intensity of the medium in the top chamber versus that in the bottom chamber (Figure 3B). These 2D airway organoids also contain abundant ciliated cells (Figure 3C). Ciliated cells were labeled by anti-β-Tubulin IV antibody (ACCTUB) and goat anti-mouse 488 secondary antibody. Confocal images were acquired using a confocal microscope. Multi-channel images were acquired using the lasers 405 nm for DAPI, 488 nm for ACCTUB, and 633/640 nm for Phalloidin. Imaging parameters were adjusted according to the user manual of the confocal microscope. Briefly, the pinhole size was set to 1 AU, the master gain was set to 650 V to 750 V with digital gain of 1.0, and the laser power was adjusted for each channel within the range of 0.2% to 5%. Image processing was performed by using the analysis software provided.
The human respiratory tract is lined with two distinct types of epitheliums, i.e., airway epithelium and alveolar epithelium. The former lines the airways from the nasal cavity to the terminal bronchiole and consists of four major types of epithelial cell, i.e., ciliated cell, goblet cell, club cell, and basal cell. In addition, the airway epithelium lining the proximal and distal airways shows a variable cellular composition along the proximal-distal axis. The proximal airway epithelium is pseudostratified, consisting of abundant ciliated cells and mucus-secreting goblet cells; whereas the distal airway epithelium is a single layer of cuboidal ciliated and club cells with less basal and goblet cells19. Human lung tissues used for deriving lung organoids have been procured from patients who underwent surgical resections due to various diseases. We use normal lung tissues adjacent to the diseased tissues for organoid culture. These lung tissues typically contain bronchioles of variable size surrounded by alveolar sacs. During the initial culture, airway epithelial stem cells or airway progenitor cells in the lung tissues survive and proliferate due to the niche factors in the expansion medium. The expansion medium enables initial derivation and long-term expansion of lung organoids by directing the organoids toward an immature state, while the airway differentiation protocol generates airway organoids phenocopying the native airway epithelium morphologically and functionally. The model system, including derivation, expansion, and differentiation of lung organoids, is outlined in Figure 4. The cellular composition in the proximal and distal airway epithelium is also illustrated in Figure 4.
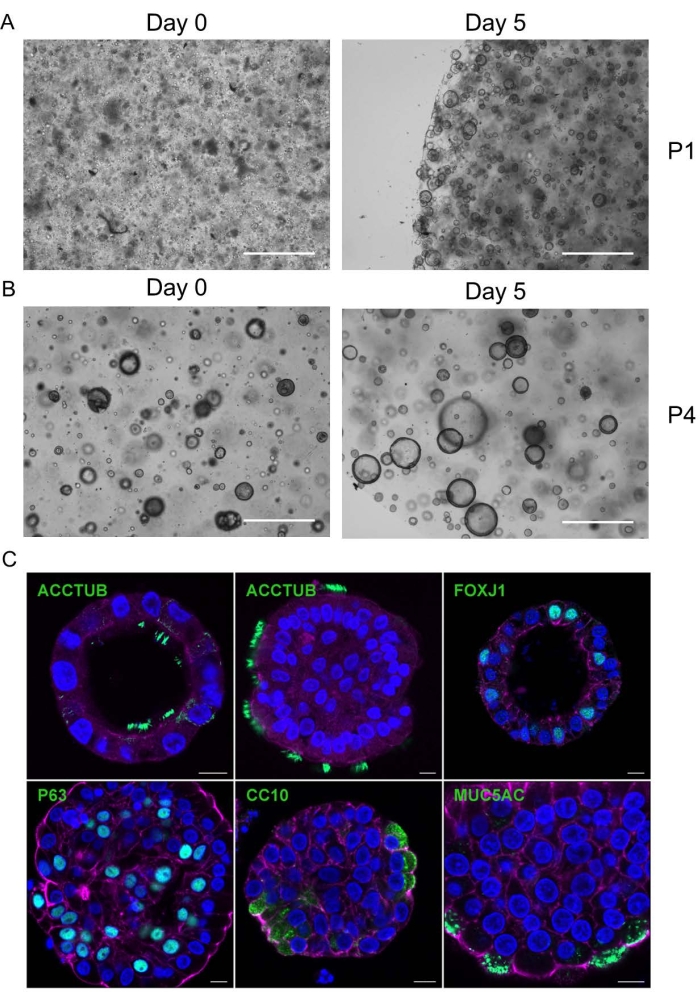
Figure 1: Derivation, expansion, and characterization of human lung organoids. (A) A representative microphotograph shows single cells embedded in the basement matrix after isolation from lung tissues on day 0 (left). On day 5, cystic organoids are growing (right). Scale bar is 0.5 mm. (B) A representative microphotograph of lung organoids on the day of the fourth passage and day 5 after passage. Scale bar is 0.5 mm. P1 and P4 represent the first and fourth passages. The images were taken at 10x magnification. (C) Confocal images of four airway epithelial cell types in human lung organoids. Four lineages of airway epithelial cells are present in the lung organoids, including ACCUB+ and FOXJ1+ ciliated cells, P63+ basal cells, CC10+ club cells, and MUC5AC+ goblet cells. Nuclei and cellular actin filaments are counterstained with DAPI (blue) and Phalloidin-647 (purple), respectively. Scale bar is 10 µm. This figure has been adopted from13. Please click here to view a larger version of this figure.
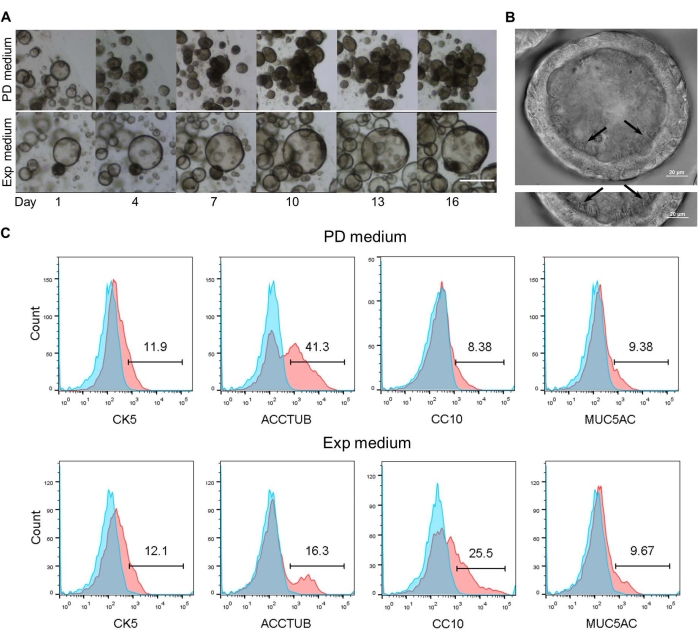
Figure 2: Proximal differentiation of human lung organoids. (A) Human lung organoids were cultured in the PD medium or the expansion (Exp) medium in parallel for 16 days. Bright-field microphotographs of organoids at the indicated days are shown. Scale bar is 0.4 mm. (B) Cilia in the differentiated airway organoids are shown (black arrow). Scale bar is 20 µm. (C) The percentages of individual cell types in organoids incubated in PD medium (top) and expansion medium (bottom) as detected by FACS analysis. The representative histograms of one organoid line are shown. The experiment was performed in three different organoid lines. This figure has been adopted from13. Please click here to view a larger version of this figure.
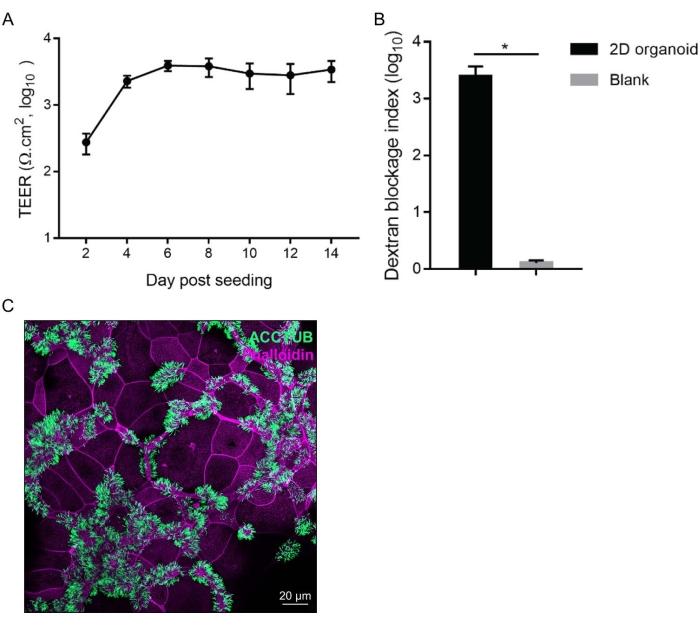
Figure 3: Generation of 2D differentiated airway organoids. (A) Trans-epithelial electronic resistance (TEER) was measured at the indicated day after incubation in the PD medium. Data show mean ± standard deviation (SD) of 2D monolayers in 10 inserts. (B) On day 10 after culture in permeable support plates, fluorescein isothiocyanate-dextran was added and the media in the top and bottom chambers were harvested for a fluorescence assay after 4 h. Dextran blockage index refers to the fluorescence intensity of the medium in the top chamber versus that in the bottom chamber. The diameter of permeable support inserts used in our experiment is 0.4 µm. Without seeding any cells, the dextran can freely penetrate the normal 2D inserts. Thus, the dextran blockage index of a normal 2D (the bar labeled with Blank) should be 1. Data represent the mean ± SD of 10 inserts seeded with 2D airway organoids (2D organoid) and those in two blank inserts (blank). (C) Confocal images of abundant ACCTUB+ ciliated cells (green) in 2D airway organoids. Cellular actin filaments are counterstained with Phalloidin-647 (purple). Scale bar is 20 µm. This figure has been adopted from13. Please click here to view a larger version of this figure.
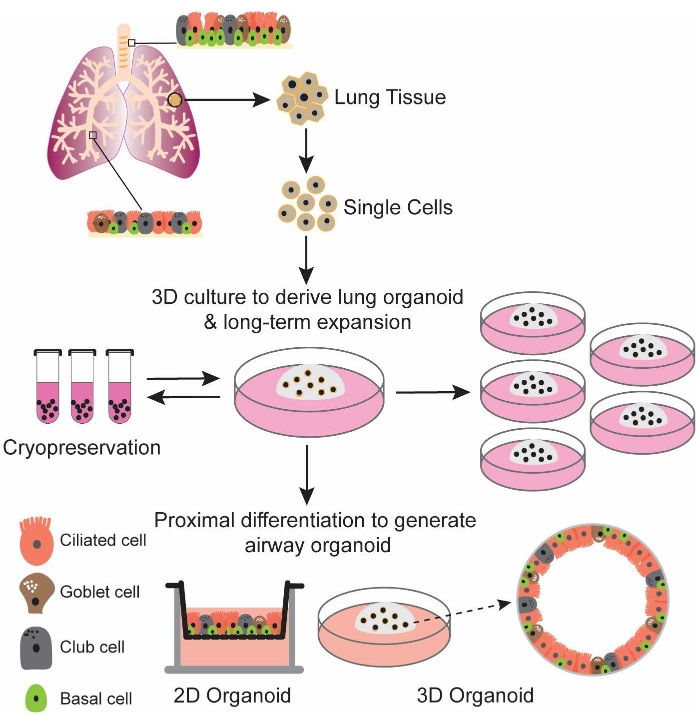
Figure 4: Schematic illustration of derivation, expansion, and differentiation of human lung organoids. Single cells isolated from human lung tissues are directly embedded in the basement matrix and incubated in the lung organoid expansion medium. The derived human lung organoids can be long-term expanded with high stability and readily recovered from cryopreserved stocks. Upon differentiation, the generated airway organoids can faithfully simulate human airway epithelium. 2D and 3D airway organoids have been developed for various experimental manipulations. Please click here to view a larger version of this figure.
Supplementary Video 1. The synchronously beating cilia drive the cell debris to swirl unidirectionally in the differentiated airway organoids13. This video has been adopted from13. Please click here to download this Video.
