Virus preparation
An example of virus purification by density gradient centrifugation is shown in Figure 1A. The light pink band, found at the interface of the loaded cellular material and the 1.25 g/mL CsCl layer, is primarily composed of disrupted cells and their debris (see magenta arrow in Figure 1A). It derives its pinkish color from the small amount of culture medium that is carried over from step 1.5 in the protocol. The virus particles of interest, which appear as a milky white band, are found at the interface of the 1.25 g/mL CsCl and 1.40 g/mL CsCl solutions (see yellow arrow in Figure 1A). One may also observe a band of material that floats 2-3 mm above the enriched virus particles (see thin black arrow in Figure 1A). It comprises unassembled viruses and debris, contains few IVP, and should be avoided when collecting the virus sample.
Further purification of the virus and buffer exchange using a PD10 column is depicted in Figure 1B, and the OD260 readings of the resulting fractions after elution are shown in Figure 1C. The void volume of these columns is approximately 3 mL, and thus the virus begins to appear in fraction 6 and peaks in fraction 9. In this experiment, fractions 6-9 were pooled. While fraction 6 and 7 are relatively low in virus particles, they contain sufficient virus to merit recovery, and they serve to dilute the very high titer fractions to a more reasonable concentration. Fractions 10-12 were not included as the increase in OD260 in fraction 11 indicates the possible presence of a second contaminating peak, which is variably observed in these preparations. The pooled fraction will typically have 1 x 107 to 1 x 108 IVP/µL, and the expected yield would be in the order of 1 x 1010 to 2 x 1011 total IVP. This is sufficient amount of virus to perform hundreds of transductions. While it is possible to titer the virus by plaque assays or by counting colonies of cells expressing fluorescent proteins22, the 1% rule is sufficient in most cases. This rule states that 1% of purified virus particles, estimated by measuring the OD260 of the sample, are IVP. Aliquots of virus stored at -80 ˚C have a shelf-life of 2-5 years, although infectivity decreases over the long term. Thawed aliquots of virus can be refrozen at -80 °C one time without significant loss of infectivity. However, repeated thawing and freezing negatively impacts viral infectivity.
Bladder transduction
An important first step when assessing the impact of transduction is to confirm transgene expression. This can be assessed using several techniques, including tools that detect mRNA (e.g., RNAScope), western blot analysis, or use of immunofluorescence9,10,23. Figure 2 is an example of mouse urothelium that was transduced with an adenovirus that encodes V5-epitope tagged human growth hormone (V5-hGH)23. This protein is packaged into discoidal/fusiform vesicles and can be exocytosed during bladder filling19,23. Western blot analysis of urothelial lysates revealed V5-hGH expression in the urothelium of transduced bladders, but not in untransduced ones (Figure 2A). Expression was also confirmed by immunofluorescence, in this case using antibodies that recognized hGH or the V5 epitope tag (untransduced bladders lacked signal, not shown) (Figure 2B).
An additional example is transduction of the rat bladder with a virus encoding CLDN2, a pore-forming tight junction-associated protein24,25. CLDN2 increases paracellular flux of cations (including K+) and its overexpression results in inflammation and development of visceral pain10. Western blot analysis confirmed expression of CLND2 in rat bladders transduced with adenovirus encoding Cldn2, but not those transduced with a control GFP-encoding virus (Figure 2C). In general, GFP is not considered to be toxic when expressed in cells and it thus serves as a useful control. The use of GFP also allows the investigator to confirm that transduction is working. Immunofluorescence analysis further confirmed exogenous CLDN2 expression in urothelial umbrella cells transduced with virus encoding Cldn2 cDNA (red signal in Figure 2D). Furthermore, similar to endogenous CLDN2 (not shown), the expressed CLDN2 is localized to TJP1-labeled tight junctions, as well as the basolateral surfaces of the umbrella cells (CLDN2 is labeled green in Figure 2E)10. In the case of CLDN2 expression, it was highest one day after transduction, but then trailed off, and was barely detectable after 15 days.
A second consideration is, which cell types will be targeted by adenoviral transduction. While in rats it is possible to mostly transduce the umbrella cell layer19, in mouse, all the layers of the urothelium can be transduced, although transduction of the intermediate and basal cell layers can be variable. Importantly, in the entirety of the bladder wall, only the urothelium is transduced and no other tissue is targeted by the instilled adenovirus (Figure 3).
While analyses of transduced cells can be performed at the single cell level, when exploring an overall bladder phenotype, one must transduce the majority of the urothelial cells. Thus, it is important to define the efficiency of transduction (i.e., what fraction of urothelial cells are transduced). For example, in the case of the Cldn2-expressing adenovirus, >95% of umbrella cells were transduced (see Figure 2D). An additional example is the image field shown in Figure 3, where counting the number of transduced umbrella cells (in this case, expressing the Ca2+ sensor GCAMP5G), reveals an efficiency that approaches 95%. However, one must examine cells in random fields taken throughout the bladder wall to achieve an accurate and unbiased estimate of the transduction efficiency.
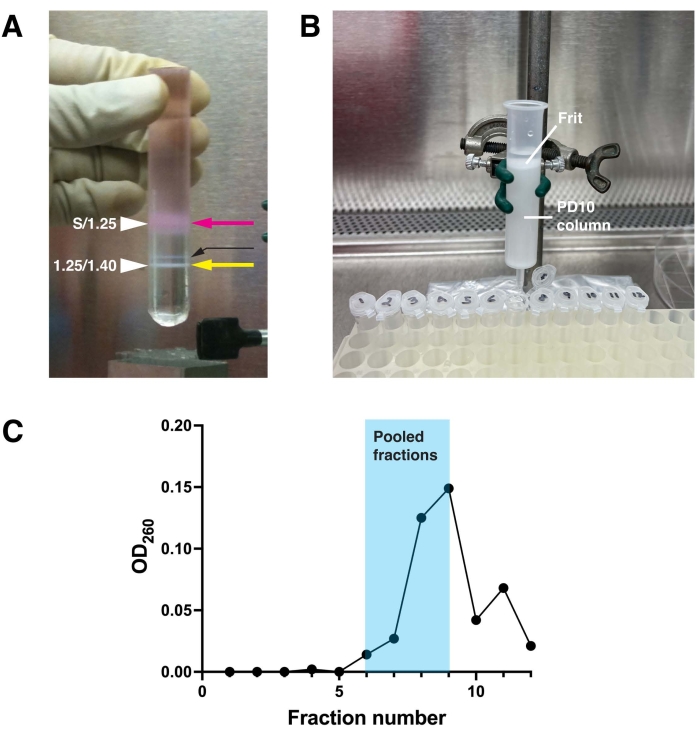
Figure 1: Purification of adenovirus. (A) Adenovirus particles produced by infected HEK293T cells were purified by centrifugation on a discontinuous CsCl gradient made up of a layer of 1.4 g/mL CsCl, a layer of 1.25 g/mL CsCl, and a layer of sample (S) diluted in Tris-EDTA solution. The cellular material (pink arrow) accumulates at the S/1.25 interface, while the purified adenovirus floats at the 1.25/1.4 interface (yellow arrow). A small band of improperly assembled virus floats above the latter (thin black arrow). (B) The adenovirus-rich band in the gradient is recovered using a needle, and the CsCl is removed from the adenovirus by gel filtration using a G25 Sephadex-filled PD10 column equilibrated with PBS containing 10.0% (v/v) glycerol. The surface of the Sephadex is protected by a porous, plastic frit. (C) Fractions, 0.5 mL, were collected from the PD10 column. The OD260 of the fractions were measured in a spectrophotometer and the values plotted. Virus-rich fractions elute in the void volume, which begins at fraction 6 and extends to fraction 9. Pooled fractions for this representative experiment are shaded in blue. Please click here to view a larger version of this figure.
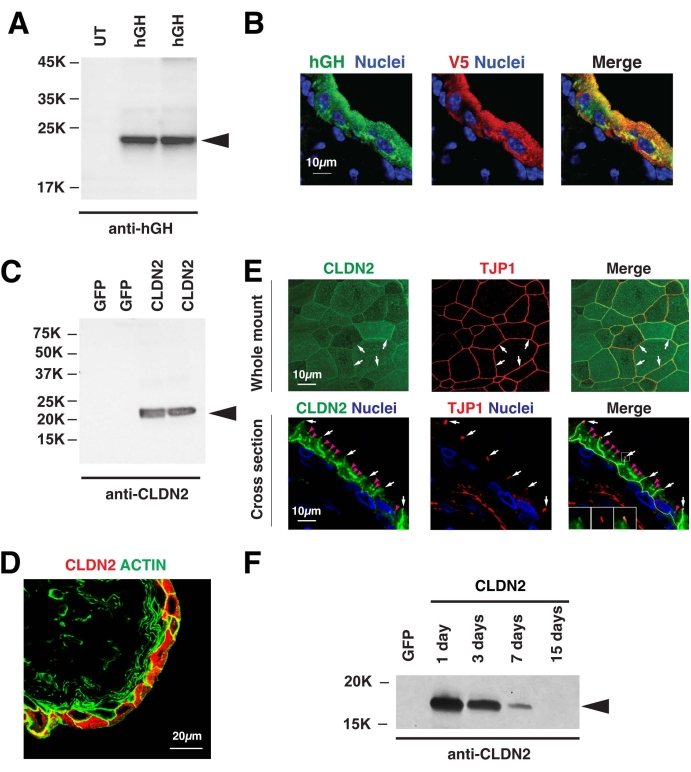
Figure 2: Transduction of rodent bladder urothelium with adenoviruses encoding V5-hGH or CLDN2. (A) Mouse urothelium was left untransduced (UT) or transduced with adenoviruses encoding V5 epitope tagged human growth hormone (hGH). After 24 h, the bladders were recovered, urothelial lysates were prepared and subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis, and V5-hGH expression confirmed using western blots probed with an antibody against hGH. (B) Detection of V5-hGH in mouse urothelium sectioned and stained using antibodies to hGH (green) or the V5 epitope (red) and fluorophore-tagged secondary antibodies. Immunofluorescence was captured using confocal microscopy. Samples were counterstained with TO-PRO3 to label nuclei. (C–F) Rat urothelium was transduced with a virus encoding rat CLDN2 (common name claudin-2) or GFP (common name green fluorescent protein) as a control. (C) Detection of exogenous CLDN2 by western blotting using an antibody again CLDN2. Note that the endogenous CLDN2 is expressed at low levels and is not detected in this experiment. (D) Transduction of the umbrella cell layer is revealed by immunofluorescence and confocal microscopy. The borders of the cell are revealed by co-staining the tissue with FITC-phalloidin, which labels the cortical actin cytoskeleton. (E) Detection of exogenous CLDN2 and TJP1 (common name ZO1) by immunofluorescence and confocal microscopy of whole-mounted or cross-sectioned urothelium. Small white arrows indicate the location of the tight junction and small magenta arrowheads mark the location of intracellular accumulations of CLDN2 in the cell cytoplasm. These were previously revealed to be Golgi-associated CLDN210. (F) Following transduction, the animals were euthanized on the indicated days post infection. Exogenous CLDN2 expression was detected using western blotting. Animals expressing GFP were euthanized after day 1. The data in Figure 2C–E have been modified from Montalbetti et al.10, and are reproduced with permission of the American Physiological Society. Please click here to view a larger version of this figure.
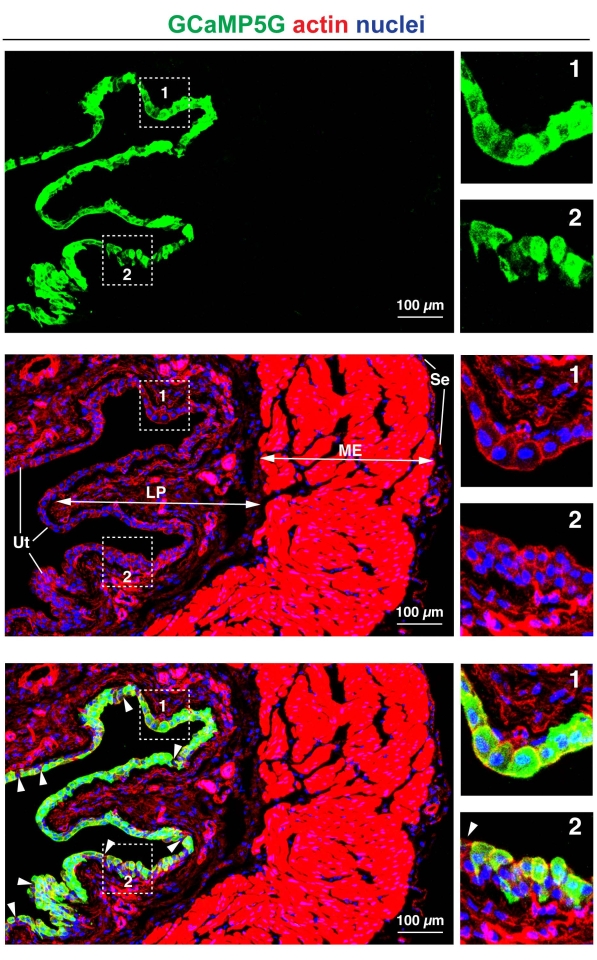
Figure 3: Efficiency of adenoviral transduction. Mouse bladder urothelium was transduced with an adenovirus encoding the calcium sensor GCaMP5G. The upper panels show staining for GCaMP5G (detected using an antibody to green fluorescent protein; GFP, green), the middle panels show the distribution of DAPI-stained nuclei (blue) and rhodamine-phalloidin-labeled actin (red), and the bottom panels are a merger of the three signals. The white arrowheads in the lower panel are the rare umbrella cells that are not transduced. The overall efficiency of transduction of umbrella cells in this image is ~95%. Note that only the urothelium is transduced. The boxed regions are magnified in the panels to the right. The region in box 1 primarily comprises transduced umbrella cells. The region in box 2 is urothelium showing efficient transduction of umbrella cells, but less efficient transduction of the underlying cell layers. LP = lamina propria; ME = muscularis externa; Se = serosa; Ut = urothelium. Images were acquired using a confocal microscope (see Table of Materials). Please click here to view a larger version of this figure.
