Various stomatal transgenic plants and mutants known to have less or more stomatal density and clustering (epf22,5, epf1 epf22,5, tmm12, a STOMAGEN-silenced line4, and transgenic lines carrying the estradiol-inducible Est::EPF1 or Est::EPF2 overexpression construct7) were used to demonstrate the effectiveness of the two phenotypic analyses presented here, which aimed to identify and characterize the genes that have a role in stomatal development and patterning. In order to produce high-quality epidermis images without the need for epidermal peels to quantify the different types of epidermal cells for analysis, it is critical to pre-adjust the sample staining time with TBO for each genotype, as each one may have different epidermal phenotypes compared to that of the wild-type control (Col-0) (Figure 1A and Figure 2A). Based on experience, genotypes with fewer stomata require longer staining times with TBO (Figure 1B and Figure 2D,E), while genotypes with more stomata and clustering require shorter staining times (Figure 1C and Figure 2B,C). As varying amounts of properly folded peptides in the total peptide solution can mask the biological activity of the peptides in stomatal development, it is good practice to use additional, higher total concentrations of the peptide solution (e.g., 2.5 μM EPF2), as well as genotypes that have less or more stomatal phenotypes (e.g., epf2), for the bioassays. The activity of EPF2 peptides, which have a role in inhibiting stomatal initiation, was easily detected by using epf2 mutants with more epidermal cells than Col-0, even if certain batches of the prepared EPF2 peptides had smaller amounts of the bioactive forms of the peptide (e.g., EPF2 peptide solution batch 1 in Figure 4).
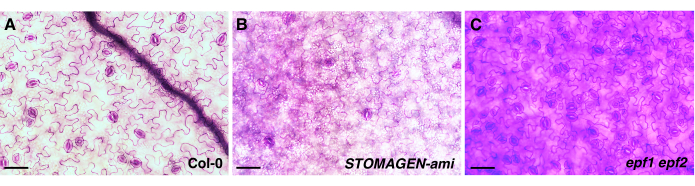
Figure 1: The effect of the incubation time with TBO on the genotypes exhibiting different epidermal phenotypes. Abaxial cotyledon images from 10 day old seedlings of three Arabidopsis genotypes: (A) wild-type (Col-0), (B) a STOMAGEN-silenced line (STOMAGEN-ami), and (C) epf1 epf2 mutants. Col-0 represents the wild-type Arabidopsis control, and the STOMAGEN-silenced line and the epf1 epf2 mutants represent the genotypes with low and high numbers of stomata, respectively. The images were taken using an inverted microscope with a 20x objective lens (0.35 mm2 field of view). The quality of the images was variable, although all the cotyledon samples from the different genotypes were stained with TBO (0.5% TBO in H2O) for the same amount of time (2 min) before imaging. Therefore, to obtain well-stained images for analysis, it is important to predetermine the adequate TBO staining time for each genotype. Scale bar = 50 µm. Please click here to view a larger version of this figure.
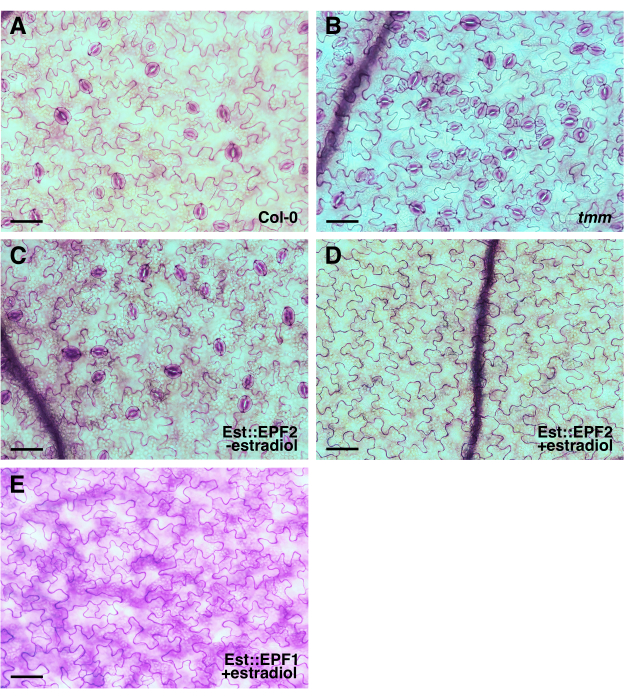
Figure 2: TBO-stained epidermis images without the use of epidermal peels for quantitative analysis. Representative epidermis images of cotyledons from 10 day old seedlings of (A) wild-type (Col-0), (B) tmm, and (C,D) transgenic lines carrying an estradiol-inducible EPF2 (Est::EPF2) or (E) EPF1 overexpression construct (Est::EPF1) can be used for the quantitative analysis of the epidermal phenotype.A cotyledon in a fixing solution for over 3 years was used to take the image presented in (E). The images were taken using an inverted microscope with a 20x objective lens (0.35 mm2 field of view). The cells were outlined by TBO staining, and half the width of the full-size images is presented for display. Scale bar = 50 µm. Please click here to view a larger version of this figure.
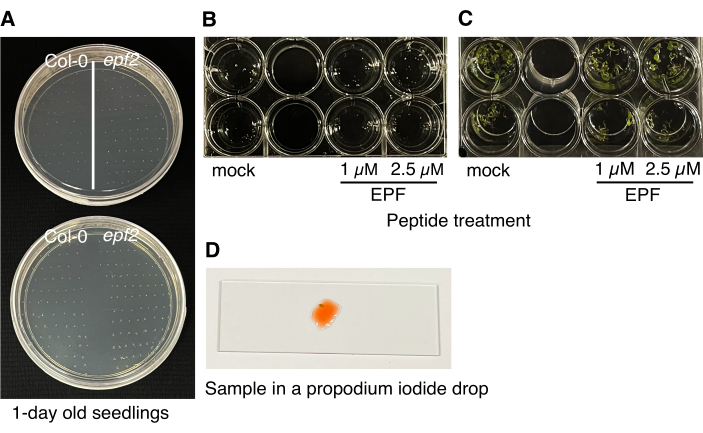
Figure 3: Procedure for the bioassays. (A) The seeds are sown on two 1/2 MS agar plates and taken out at 10 h intervals. (B) The 1 day old Arabidopsis seedlings are transplanted to 24-well plates containing 1/2 MS medium with either mock or two different peptide concentrations. (C) After 5-7 days, the seedlings are ready for imaging to determine the biological activity of the peptides in stomatal development and patterning. (D) A cotyledon slice is placed in a drop of propidium iodide solution on a microscope slide for imaging. Please click here to view a larger version of this figure.

Figure 4: Confocal microscopy of the abaxial epidermis of Arabidopsis cotyledons following peptide treatment. Representative confocal images of (A) wild-type and (B) epf2 cotyledon epidermis grown for 6-7 days in a buffer solution and an (C–F) epf2 cotyledon epidermis grown with two different batches of EPF2 peptides. The images were taken using a confocal microscope using a 40x objective lens (excitation of 561 nm and a 561 nm long-pass emission filter) to capture the propidium iodide staining and visualize the cell outlines. Each EPF2 peptide solution prepared has different amounts of properly folded (bioactive) and misfolded (inactive) forms of the peptides. Therefore, for the initial screening of peptides with a potential role in stomatal development, it is recommended to use two different concentrations of the total peptide solutions as indicated for the bioassays. Scale bar = 30 µm. Please click here to view a larger version of this figure.
