1. Protocol Design
- Calibration: Before a set of experiments, calibrate the loudspeakers. This is important so that loudspeaker display the exact volume that was set by the experimenter. Also calibrate the sensitivity of the transducer platform of the startle boxes according to the supplier’s manual. The transducer converts the vertical movement of the platform into a voltage signal. Make sure that there are no ongoing experiments when calibrating the system, and that all boxes are calibrated the same way.
- i/o function: If new strains of mice or rats are measured, an input/output function should be established. After an acclimation period of 5-10 minutes with a constant background white noise of 65 to 68 dB (see below), startle stimuli (20 ms white noise) should be displayed every 20 sec, starting at around 70-75 dB. Startle stimulus intensity will be increased between each stimulus by 2-5 dB until reaching 120-130 dB, resulting in 10-30 trials with startle stimuli (see figure 1).
- Protocol structure: Habituation and prepulse inhibition can be measured within one protocol. The protocol is divided into an acclimation period, a block I (habituation), immediately followed by a block II (PPI, figure 2). Before measuring prepulse inhibition, animals should always undergo startle habituation, so that startle attenuations due to habituation do not interfere with PPI measurements.
- Acclimation period: Each time an animal is tested, it first undergoes an acclimation phase in order to adapt to the animal holder, startle box and background noise. During a 5-10 minutes acclimation period, the constant background noise of 65-68dB white noise (depending on the noise of the environment) is displayed, but no startle stimuli. During this phase the animal will calm down, stop to explore the environment and stop moving around.
- Block I habituation: For short-term habituation (STH), between 30 -100 startle stimuli should be applied on the background. Startle stimuli are commonly white noises of 20 ms duration and very steep rise times (0, if possible). The intensity is ideally at the volume where the i/o function reached the plateau of maximum startle response, commonly at 105 to 115 dB. The intervals between single trials should be either always 20 sec or randomized between 10 and 30 sec (see discussion and figure 3).
- Block II prepulse inhibition: In order to measure PPI, trials with a startle pulse alone and trials with a prepulse are pseudorandomized in block II. Background noise and startle stimulus are the same as in block I. The prepulse is a white noise of 4 ms duration and also steep rise time. Two parameters can be varied: the interstimulus interval between prepulse and startle pulse and the intensity of the prepulse (see discussion). We propose to commonly use two different prepulse intensities (75 and 85 dB) and two different interstimulus intervals (30 ms and 100 ms). Thus there are four different prepulse-pulse trials plus the startle pulse alone trials to be pseudorandomized and displayed 10 times each, which totals 50 trials. Inter-trial intervals can be either 20 sec or randomized between 10 and 30 sec (see discussion). In some cases it might be beneficial to add a sixth type of trials, which is a prepulse alone trial (see discussion and figure 4).
- Long-term habituation: In order to measure LTH, the entire protocol is run on at least five subsequent days. Alternatively, only the acclimation phase and block I could be run, however, in order to see LTH, block I should contain at least startle 100 stimuli. The presentation of 30 stimuli per day leads to very little or no LTH in most animals, especially in mice. Runs should be on approximately the same time of each day, since startle response amplitudes fluctuate with the diurnal cycle.
2. Handling and Acclimation of Animals
There are big differences in the handling and acclimation of rats versus mice. Mice will be placed into the appropriate animal holder (they should not be restrained) for 2-5 minutes with background noise but no startle stimuli (acclimation phase of the program). This procedure should be repeated 3-5 times, once or twice a day, until defecation and urination in the mouse holder ceases or considerably decreases. Animal holders should be always replaced or cleaned after an animal is removed.
Rats should be handled for at least three sessions17. At the end of the third handling sessions they are placed into an appropriate animal holder (no restrain) and exposed to background noise for several minutes. After removing them, they can be rewarded with sunflower seeds in order to form positive associations with the testing procedure. This procedure is repeated two more times, gradually expanding the acclimation time, before the entire protocol is run.
For testing sessions, animals are placed into all chambers, doors closed and the protocol with acclimation phase, block I and block II is run. If there are different groups of animals (injections, genotypes), they should be mixed or randomized over the different runs and the different boxes. If an animal is repeatedly tested (e.g. with different treatments), it should be re-tested in the same box. For repeated PPI testing in rats, we also recommend to run an entire protocol before the actual data collection takes place. PPI often improves between the first and the second testing session (PPI learning), and stays consistent thereafter. It will also eliminate a big portion of LTH.
3. Data Analysis
- Short-term habituation: For short-term habituation analysis, all startle responses of block I are plotted for each animal. If animals within a group have similar startle response amplitudes, values can be averaged between animals. In most cases, however, absolute startle amplitudes differ considerably between animals and startle levels are not normally distributed. In this case it is more viable to normalize the data of each animal to its first, or the average of the first two, startle responses in block I (animals sometimes fall asleep during the acclimation phase resulting in a low first startle response and a high second startle). The normalized data can then be averaged across all animals in order to plot the course of habituation. For a quantitative assessment of the amount of habituation, a score can be calculated for each animal, e.g. the average of the last 10 startle responses divided by the average of the first two responses (figure 6).
- Prepulse inhibition: For analyzing prepulse inhibition, the data of block II has to be sorted according to the type of trial (e.g. by exporting all relevant data columns into excel and sort by prepulse intensity and duration of ISI). The ten traces per trial type are then averaged, and the resulting values for the prepulse-pulse trials are divided by the startle pulse alone value and multiplied by 100. This reveals the amount of remaining startle (in percent of baseline startle) under different prepulse conditions for each animal. Baseline startle (pulse alone) is 100%. These values can then be averaged across animals of a group and be plotted (figure 7a). Alternatively, the amount of PPI can be plotted by subtracting the remaining startle response from 100% (figure 7b). Please be aware: when you compare PPI in different groups of animals, you should always also report whether there is a difference in baseline startle amplitudes, by e.g. comparing the absolute startle amplitudes of the startle pulse alone trials (or startle amplitudes in block I).
- Long-term habituation: In order to analyze LTH the first two responses of block I of each day are averaged and plotted over the minimum of five consecutive testing sessions. This eliminates the possibility that differences in STH affect the result of LTH analysis. If it has been established that a treatment/gene does not affect STH, alternatively all responses in block I can be simply averaged for each day and be plotted. LTH can be quantified by calculation of a habituation score where the last days’ value is divided by the first days’ value and multiplied by 100, so that the percentage of initial startle level remaining after LTH is displayed. Habituation scores can then be averaged across animals (figure 8).
4. Representative Results:
- i/o function: Rodents typically begin to startle from a volume of 85-90 dB on (with 20 ms duration, white noise). The startle response increases with increasing volume and normally reaches a maximum at 100-110 dB. If animals deviate considerably from these values, animals might have disrupted hearing abilities or motor abilities. Typical i/o functions are displayed in figure 5.
- Short-term habituation: Well handled rats normally habituate to around 60% of their initial startle response; however, there are huge individual differences and also strain differences. The strongest habituation effect occurs normally within the first several stimuli. Mice do generally habituate less than rats (typically to about 80%), but strain differences can be very large. A typical habituation course is shown in figure 6.
- Prepulse inhibition: Most rats show PPI of around 90% with an optimal prepulse (85dB, 4 ms, white noise). PPI is very robust and individual differences are relatively small with these experimental settings. Lower volume prepulses yield less PPI and more variability (even within an animal), but also seems to be more vulnerable to pharmacological or genetic manipulations. Different PPI results are plotted in figure 7.
- Long-term habituation: Long-term habituation can be observed over several testing sessions. LTH is very robust in rats. In mice, it often requires the presentation of a lot of startle stimuli in each session in order to observe LTH. Typical LTH results can be seen in figure 8.
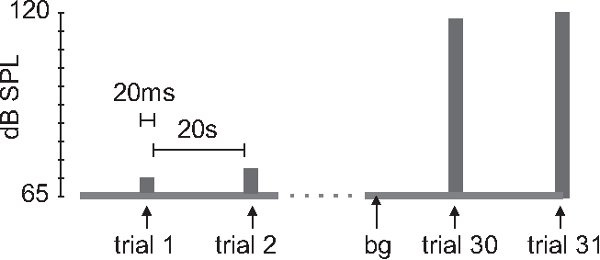
Figure 1. Stimulus protocol for i/o function. After an acclimation period of 5-10 min. with 65 dB sound pressure level (SPL) background noise and no startle stimuli (not shown), 20 ms white noise stimuli are presented every 20 sec. The intensity gradually increases from 75 to 130 dB in 5 dB increments (bg = background noise).

Figure 2. Protocol structure for combined habituation and PPI measurement. During the whole protocol, a constant background noise of 65 dB is applied. There is an acclimation period of 5-10 min. without any further stimulation. Immediately thereafter, habituation is tested by 30-100 startle stimuli (block I, see figure 3). This is immediately followed by PPI testing (block II, see figure 4).
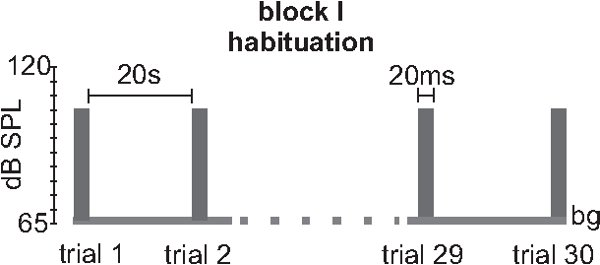
Figure 3. Stimulus protocol for measuring habituation (block I). An example for a typical block I for testing short-term habituation is shown. It consists of 30 100 identical trials where a 20 ms 105 dB white noise with a 0 rise time is presented with an inter-trial interval (ITI) of 20 sec. Variations of this protocol may include higher startle stimulus intensities or variable ITIs
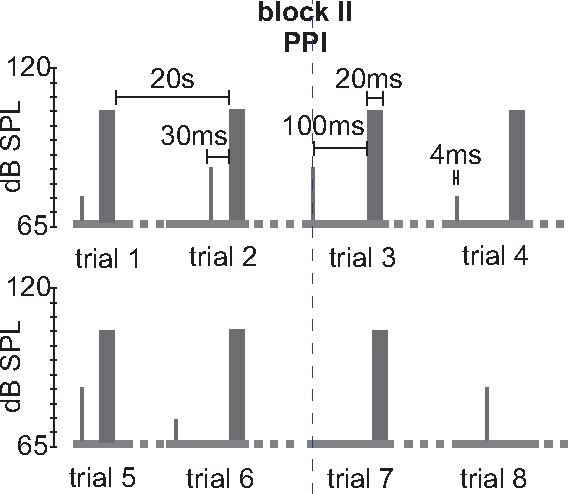
Figure 4. Stimulus protocol for measuring PPI (block II). An example for a typical part of a block II for testing PPI is shown. Block II consists of 5-6 different trial types that are presented 10 times each in a pseudorandomized order. Here, two different prepulse intensities (75 dB and 85 dB) and two different interstimulus intervals (ISIs, 30 and 100 ms) are tested. Startle stimulus alone trials and prepulse alone trials are interspersed. This block would have 6×10= 60 trials. Prepulses are 4 ms white noise pulses with 0 rise time. Variations of this protocol would consist in variable ITIs, higher startle stimulus intensities, different prepulse intensities and/or durations, and different ISIs between prepulse and pulse.
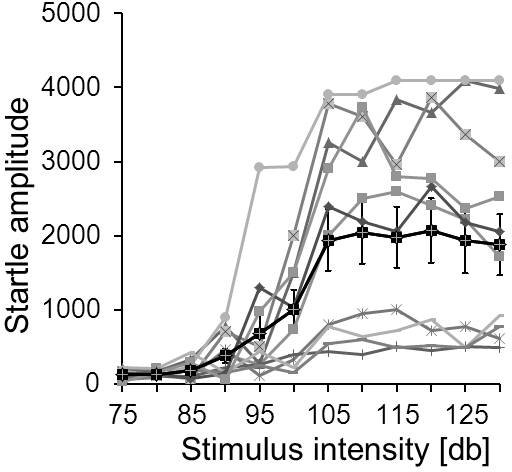
Figure 5. Example for an i/o function. The input/output curves of 11 individual mice of the same strain are displayed in grey. In this case, the individual startle amplitudes vary considerably (startle responses are in arbitrary units). The solid black line shows the average startle amplitudes and standard errors at different startle stimulus intensities. These mice reached their maximum startle response at around 105 dB.
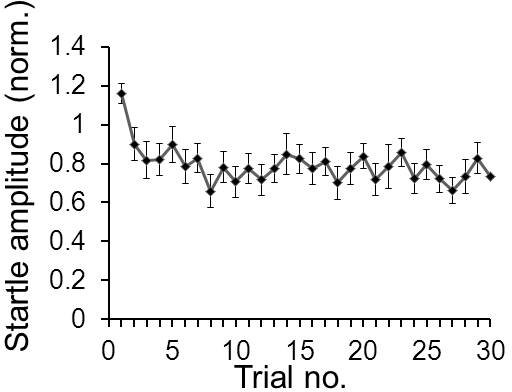
Figure 6. Example for short-term habituation data. A typical average short-term habituation curve of 20 mice is shown. Startle amplitudes of each mouse in response to 30 startle stimuli were normalized to the average of its first two startle responses in trials 1 & 2. The normalized data was then averaged across mice and the standard error was calculated.
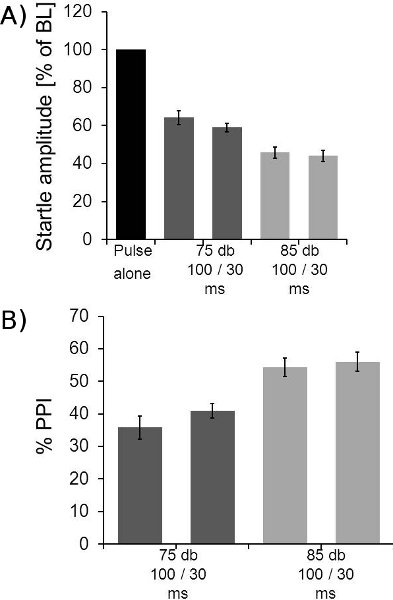
Figure 7. Example for PPI data. A: Averaged PPI data of 8 mice is shown. The 10 startle alone trials of block II were averaged for each mouse and the averages of the other trial types expressed as the percentage of the stimulus alone startle amplitudes. The figure shows the startle response amplitudes under different prepulse conditions. Two different ISIs (30 and 100 ms) and two different prepulse intensities (75 and 85 dB) were measured. B: Same data as in A, but plotted as amount of PPI in percent of baseline startle. Data shown above was subtracted from 100. These mice showed a maximum PPI of around 50%. Please note that the same protocol yield PPI in most rat strains of around 90%.
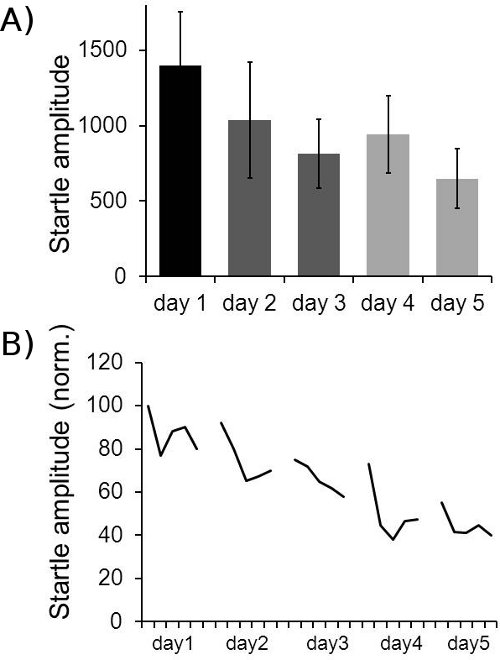
Figure 8. Example for long-term habituation data. A: Averaged LTH data for 18 mice is shown. The first two startle responses in block I of each day were averaged across all mice. The relatively large standard error bars are mainly caused by differences in absolute startle amplitude between individual mice. B: Normalized startle amplitudes of 18 mice over five days. In order to reduce noise, groups of 6 consecutive startle responses in block I (30 stimuli) were always averaged per animal, resulting in five values for block I for each animal per day. These were normalized for each animal to the first value of the first day (100%). The average over all 18 animals is displayed. It shows STH within each day, as well as LTH across five days.
