1. Expression of Toxic PolyQ-expansion Proteins in Yeast
A systematic analysis has established the precise amino acid sequence of a polyQ-expansion protein that is required to produce toxicity in yeast 7. This toxic polyQ-expansion protein contains an amino-terminal FLAG-tag followed by 17 amino acids from the original sequence of the huntingtin protein, a polyQ region, and a carboxy-terminal fusion to a fluorescent protein (either GFP or CFP, see Figure 1 a). The expression of proteins with polyQ expansions of 46 glutamines or more (e.g. 72 and 103 glutamines) produces toxicity in yeast when expressed under the control of the inducible and relative strong GAL1 promoter 7, 9.
As described previously, polyQ toxicity in yeast is only apparent in cells that carry the protein Rnq1p in its prion conformation, [RNQ+], e.g. the yeast strain W303 9. The toxic polyQ-expansion protein recapitulates central aspects of polyQ-biology in yeast, such as polyQ length-dependent toxicity (see below) and aggregation (Figure 1 b). Notably, toxic polyQ-expansion proteins do not kill yeast cells immediately; they rather impair or arrest the cell cycle and cell devision, thereby slowing down or inhibiting the growth of yeast colonies on plates or liquid cultures (our unpublished data).
2. Potential Problems with Yeast Cells Expressing Toxic PolyQ-expansion Proteins
Yeast cells expressing the otherwise toxic polyQ expansion proteins may not show any growth defect 9. The genetic nature of these suppressors of polyQ-toxicity does not appear to be based on simple Mendelian mutations and thus can occur with relatively high frequency (our unpublished results). We speculate that these spontaneous suppressors are caused by the curing of unidentified prions that function similarly to [RNQ+] in determining polyQ toxicity.These spontaneous suppressors of polyQ toxicity can jeopardize the successful outcome of any experiment that aims to characterize polyQ toxicity or to identify and characterize modifiers of polyQ toxicity.
In order to avoid the frequent occurrence of these spontaneous suppressors, follow the precautionary measures outlined below, which have proven to be very effective:
- Use fresh yeast cells for toxicity experiments. Do not store yeast for extended periods of time. Frequently retrieve fresh yeast colonies from frozen stocks.
- Frequently monitor the expression and aggregation of toxic polyQ-expansion proteins by fluorescence microscopy.
- Keep the yeast cells in media that repress the expression of the toxic polyQ toxicity at all times (i.e. in selective media containing glucose as sole carbon source) before starting any toxicity measurements.
- Use at least three independent transformants for each polyQ-toxicity experiment.
3. Growth Assays
- For each experimental condition, inoculate one colony each from three independent transformants of yeast cells harboring the toxic polyQ-expansion protein in 3 ml of selective yeast media with glucose as sole carbon source.
- Incubate these cultures overnight at 30 °C. Under these conditions, the cells are not expressing the polyQ-expansion protein because the glucose in the medium represses their expression. Do not let these cultures overgrow; keep the OD600 (absorbance of light of 600 nm wavelength) of the overnight cultures below 1.
- We routinely use three different assays to monitor the growth defects of yeast cells expressing the toxic polyQ-expansion protein:
3.1 Growth on plates
- Dilute the overnight yeast cultures (grown with 220 rpm shaking) to an OD600 of 0.0005 (i.e. a 1:1000 dilution of a OD600 = 0.5 culture) in selective media 7 containing glucose.
- Evenly spread 50 μl (resulting in ca. 700 colonies per plate) of each diluted culture on one plate (10 cm diameter) with selective medium containing glucose as sole carbon source and one plate with selective medium containing galactose as sole carbon source.
- Incubate the plates for three to four days at 30 °C. After the incubation, take photographs of each plate and count the number of colonies on the glucose and the number of colonies on the galactose plates. Under ideal conditions, there should be no or only very few colonies on the galactose plate when using yeast cells expressing a highly toxic polyQ-expansion protein (103Q, Figure 2a).
3.2 Spotting assays
This assay is more quantitative than the plating assay described above and can thus reveal even subtle differences in polyQ toxicity with the same experiment on the same plate.
- Dilute the overnight cultures grown in medium with glucose to an OD600 of 0.1.
- Pipette 200 μl of these diluted cultures into sterile 96-well plates and prepare five fivefold serial dilutions in sterile water using a multi-channel pipette.
- Using a frogger,(also called a spotter, with 8 x 6 pins for transfer of cell suspensions) transfer the cell suspensions onto plates containing selective media with glucose as sole carbon source and plates containing selective media with galactose as sole carbon source.
- Allow the plates to dry before incubating them at 30 °C for three to four days.
- After the incubation, take photographs of each plate (Figure 2b).
3.3. Monitoring polyQ toxicity in yeast by growth of liquid cultures
This protocol is the most quantitative (OD600 numbers) of the three assays described here and can even detect very subtle differences in polyQ toxicity. The aforementioned occurrence of spontaneous suppressors, however, can potentially produce misleading results. I therefore recommend combining this assay with at least one of the two plating assays described above. We propose to use the BioscreenC instrument for these experiments. The BioscreenC is an instrument that automatically measures the optical density of yeast cultures in 100-well plates while incubated at defined temperatures with defined agitation. Other methods for growing yeast cells and measuring their optical density may also apply.
- Wash the yeast cells from minimal media containing glucose as sole carbon source three times in 3 ml sterile water.
- Dilute the overnight cultures grown in medium with galactose to an OD600 of 0.1.
- Fill each well of the 100-well BioscreenC plate with 300 μl of the yeast cultures.
- Open the Easy Bioscreen Experiment program. Determine the number of samples you wish to monitor (including blanks and medium-only controls), set the temperature to 30 °C, set the length of the experiments to 3 days, set the measurement intervals to 15 minutes, set the filter to 600 nm /Brown, and set the shaking mode to 15 seconds before each measurement at medium strength.
- The BioscreenC instrument and the attached software will produce Excel spreadsheets of each data point taken during the experiment.
- Prepare growth curves with Excel for each sample and compare growth of different samples (Figure 2c). A detailed description of the analysis of data produced by BioscreenC experiments has been provided before 10.
4. Representative Results
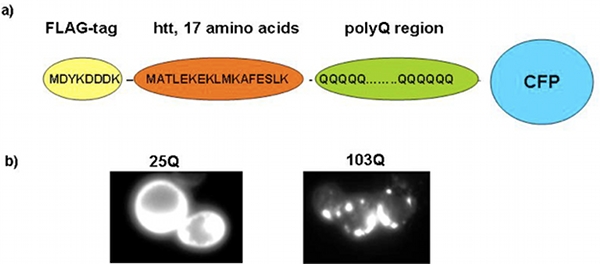
Figure 1. The yeast polyQ model. a) Schematic representation of the toxic polyQ-expansion protein. b) Fluorescence microscopy showing yeast cells expressing a short, non-toxic polyQ expansion protein (25Q, left panel) and yeast cells expressing a long, toxic polyQ-expansion protein (103Q, right panel).
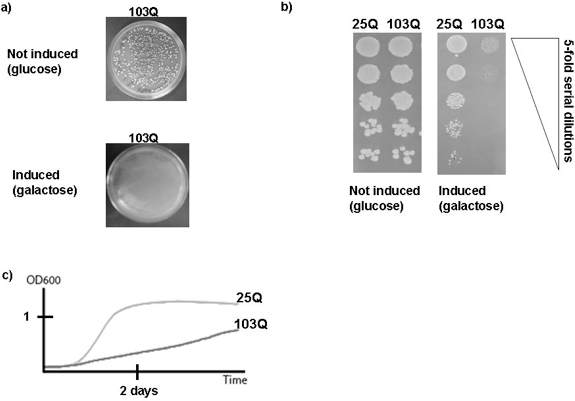
Figure 2. Representative results of growth assays of yeast cells expressing polyQ-expansion proteins. a) Plating assay. Approximately 700 yeast cells were spread on plates and incubated for three days at 30 °C. The upper panel shows a plate containing glucose medium, i.e. the expression of the toxic polyQ-expansion protein (103Q) is not induced. The lower panel shows a yeast plate containing galactose medium, i.e. the expression of the toxic polyQ-expansion protein is induced. Note that in the experiment shown here, no spontaneous suppressor occurred. b) Spotting assay. Five serial five-fold dilutions of yeast cells harboring either a non-toxic polyQ protein (25Q) or a toxic polyQ-expansion protein (103Q) were spotted on glucose plates that repress the expression of the proteins (left panel) or on galactose plates that induce their expression. The plates were then incubated for three days at 30 °C. c) BioscreenC experiment. The growth of cultures of yeast cells expressing either a non-toxic polyQ protein (25Q) or a toxic polyQ-expansion protein (103Q) were monitored by the BioscreenC instrument. The experimental conditions and the analysis of the BioscreenC experiments are described in the main text.
