Optimization of the procedures for quantifying neuromast regeneration of the lateral line in adult zebrafish.
The neuromasts of larval zebrafish are readily quantifiable; however, the lateral line of the adult zebrafish has a much greater number of neuromasts per stitch making quantitative analyses more difficult6,17,19,20. As seen in Figure 1A, the head has a significantly higher number of neuromasts compared to either the mid-section or tail; with the tail region having the least number of neuromasts as shown in Figure 1D. Because the pattern of stitches in the head is complicated and significantly greater in the number of neuromasts, it did not lend itself as a region for quantitative analysis. In addition, regardless of the gentamicin concentration we tested, complete ablation of neuromasts throughout the head was rarely attainable; leaving spots of neuromasts observed after gentamicin treatment as previously reported by Van Trump et al.17 In contrast, the tail has too few neuromasts, and as such, we selected the mid-body region (Figure 1B) to quantitatively analyze neuromast regeneration in the adult. In this region, we identified four stitches just posterior to the lateral pectoral fin that were consistent in neuromast number among all adults [61.45 (n = 95)] (Figures 1B and 1E). Importantly, we were able to consistently and completely ablate the neuromasts of this region by a 24 hr 0.004% gentamicin treatment (as previously reported in Van Trump et al.17) allowing for a subsequent accurate determination of neuromast regeneration (compare Figures 1B and 1E and inset with Figure 2, 0 hr).

Figure 1. The fluorescent pattern of neuromast within stitches of the adult zebrafish is shown along the longitudinal axis. Panel A is the head region, Panel B is the Mid-body region with the four stitches used for quantitative analysis outlined with a box. Panel C is the Posterior-body region. Panel D is the caudal fin region. A higher magnification of the 4 stitches of the Mid-body region used for quantitative analysis is shown in Panel E. Magnification Power of 1X and 2X.
It has been reported that copper sulfate treatment is an effective chemical method to induce rapid necrosis of hair cells in embryos and larvae21. Here we tested copper sulfate treatment with the hope that it might shorten the time to induce neuromast ablation. Copper sulfate concentrations ranging from 5-50 mM for various exposure times up to 48 hr were utilized as was previously reported by Liang et al.21 It was found that copper sulfate was lethal at the higher concentrations and not effective at lower concentrations in adult fish (data not shown). Please click here to view a larger version of this figure.
Parameters used for fluorescent analysis of lateral line regeneration in adult zebrafish.
Regeneration was monitored after all neuromasts within the four mid-body stitches were ablated following 24 hr gentamicin-treatment (Figure 2, compare 0 hr with control) and positive regeneration was determined by the appearance of a minimum of three neuromasts within a stitch. (Figure 2, 0 hr). By 8 hpg, approximately one third of the fish had some sign of recovery (n = 34); although the intensity of neuromasts was faint in the regenerating stitches (Figure 2, 8 hr, faint stitches outlined by boxes). The number of neuromasts and their intensity continued to increase in a linear fashion until regeneration reached a plateau at 16 hpg (compare Figure 2, 16 hr with Figure 2, 8 hr). It was not until at least 24 hpg that all fish treated with gentamicin had fully recovered with both equal numbers and intensities of the neuromasts within the lateral line stitches as compared to controls (Figure 2, 24 hr). A time-line for neuromast regeneration following gentamicin withdrawal is shown in Figure 3 which shows the linear and plateau phases of the recovery curve. We note that in less than 5% of cases, the regenerating stitches did not appear as individual entities but instead appeared as a smear of fluorescence.

Figure 2. Fluorescent images of neuromasts. Control fish, 0 Hr fish (immediately following 24 hr of 0.004% gentamicin treatment), 8 Hr fish (8 hpg) with some faint staining of neuromasts within the 4 stitches used for quantitative analysis (only 30% of all fish showed this pattern of staining at 8 hpg; 70% showed no neuromast staining at this time point; the white boxes outline the faintly stained neuromasts that were seen in the 30% of fish that showed some degree of regeneration at 8 hpg), 16 Hr fish, and 24 Hr fish. Complete regeneration of neuromasts within the 4 stitches in regard to 1) the number of neuromasts and 2) intensity of staining of neuromasts was observed by 24 hpg. Please click here to view a larger version of this figure.
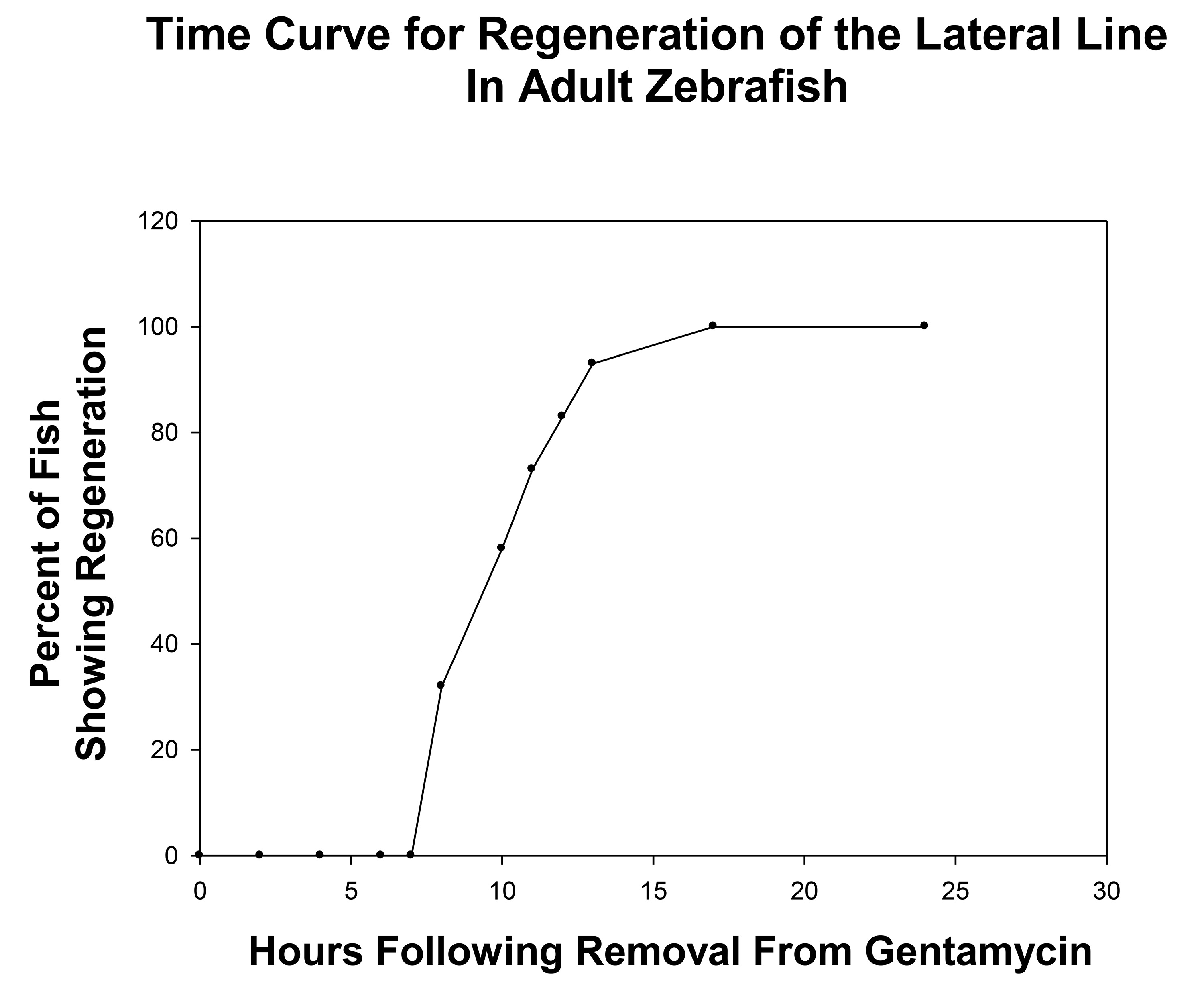
Figure 3. Graph showing the time course of neuromasts regeneration following withdrawal from 24 hr treatment of adult zebrafish with 0.004% gentamicin. As shown, recovery begins at the 8 hpg time point and reached a plateau at the 16 hpg time point. This defined the liner phase of neuromast regeneration between 8-16 hr and time points within this linear phase should be used to quantitatively compare control and experiment groups.
Neuromast toxicity is induced by prolonged exposure to fluorescent staining dyes.
In our estimation an ideal way to perform these experiments would be to stain the fish with the fluorescent dye prior to treatment, stain again at 0 hr then stain again at regeneration time points. However, we encountered a complication to these studies which is the fact that hair cell fluorescent staining dyes such as 4-Di-2-Asp, as can also be the case with other mitochondrial stains22 can have a toxic affect on hair cells23. This fact required us to use separate groups of fish since repeated staining of the same fish could not be employed. In all cases the experimental fish including controls were treated in parallel to eliminate experimental variability.
Confocal analysis at the level of individual hair cells.
If the results obtained from the neuromast analysis are not statistically significant between the control and experimental groups, one may extend these studies to the level of the individual hail cells to obtain a higher degree of resolution for quantitative comparisons. As indicated in Figure 4, neuromasts from a control group (a neuromast at 12 hr of regeneration is shown in this figure) can be viewed by confocal microscopy of skin preparations from the mid body region. At 8 hr, 10 hr, and 12 hr of regeneration time, we found that control groups (7 animals/group) had a range of 0-4 hair cells/neuromast. As expected for control groups, when quantitatively analyzed, no statistical difference between neuromasts was detected in terms of the number of hair cells per neuromast at the time points indicated above (P values ranged from 0.230-0.472). Such an approach may be taken between any control and experiment group when needed to extend or confirm the data obtained from the first phase of neuromast studies.
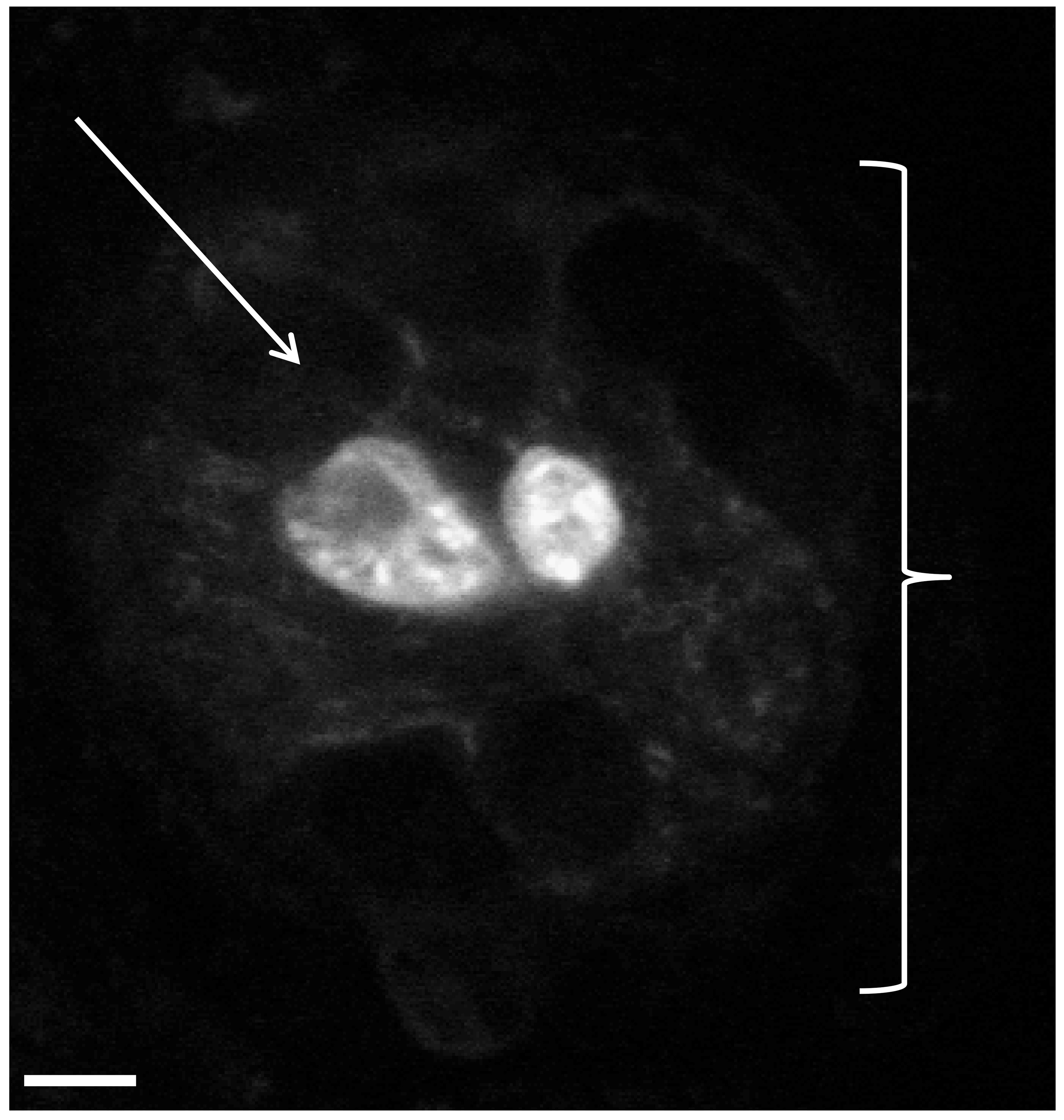
Figure 4. Analysis of hair cell/neuromast regeneration using skin preparations of the defined lateral line region described in protocol step 3.3. Fluorescent confocal image of hair cells within a neuromast obtained from a zebrafish skin preparation. Two vital dye stained hair cells are shown within a neuromast of a control fish (Figure 4). This image was obtained 12 hr post-removal of gentamicin (linear regeneration phase). The white bracket symbol ( ) indicates an individual neuromast while the white arrow indicates a supporting cell surrounding the hair cells. Supporting cells do not stain under these conditions and appear as black spaces. Magnification, 60X.
| 1 | 1. Gentamicin treatment [0.004% (4.32 mM)] of control and experimental fish for 24 hr at 28 °C using an incubator. |
| 2 | 2. Wash out of gentamicin to initiate regeneration of hair cells. Return fish to the 28 °C incubator for investigator selected time periods between 8-16 hr. |
| 2 | 3. Vital dye stain [0.08% 4-4-diethylaminostyryl-N-methylpyridinium iodide (4-Di-2-Asp)] control and experimental fish for 1 hr at room temperature and then wash out stain with fish water for fluorescent imaging. |
| 1 or 2 | 4. If necessary due to nonsignificant results from analysis of neuromasts, repeat Protocols 1-3 with a separate group of control and experimental fish, but then obtain a skin preparation for confocal analysis of individual hair cells. |
Table 1. A summary of the protocol outlined above.
