تقرير الكمي من التفاعلات الكيميائية الحيوية يوفر نظرة ثاقبة العمليات البيولوجية في أساس الحياة. الكالوري يقدم منهجية خالية من التسمية لوصف كميا كل تفاعل كيميائي تقريبا في الحل. يقيس هذا الأسلوب الحرارة المنطلقة أو الممتصة مع مرور الوقت، وبالتالي فهو نظام الكشف الشامل ومنهجية مريحة للغاية لقياس كمية من رد فعل جزيئات (أي الديناميكا الحرارية ملزمة)، وكذلك لقياس معدل التفاعل (أي حركية). على وجه الخصوص، اعتمد متساوي الكالوري المعايرة (ITC) وطريقة الاختيار لتوصيف الديناميكا الحرارية من التوازنات الجزيئية البيولوجية، التي تنطوي على البروتين يجند، البروتين البروتين، البروتين أيونات المعادن والتفاعلات الحمض النووي البروتين 1-6. بالإضافة إلى ذلك، قدرة المركز على توفير المعلومات الحركية يجعل من نظام قوي جدا لقياس انزيم الحفز، على الرغم من أن إمكانات هذا التطبيق لا يزالالتقليل 7-9.
المعادلة ميكايليس-مينتن 10 هو الوصف الكمي من التفاعلات الإنزيمية، كما أنها توفر علاقة بين معدل التفاعل وتركيز الركيزة، اعتمادا على معلمتين الحركية: ثابت ميخائيل (K M) وثابت معدل الحفاز (ك القط) . القط ك / يشار نسبة K M كما كفاءة الحفاز للانزيم. في الممارسة العملية، وتحديد K M وك القط لردة فعل معينة ويقدم وصفا كاملا للالحفز.
في رد فعل الأنزيمية نموذجي (الشكل 1)، وهي الركيزة (S) يتفاعل مع انزيم (E) تشكيل مجمع انزيم الركيزة (ES)، والتي يتم تفعيلها لاحقا في الحالة الانتقالية (ES *). يتم تحويل هذا الأخير إلى الإنزيم المنتج (EP) المعقدة التي تنأى في نهاية المطاف. هذه الخطوةموصوفة و من رد الفعل التالية.
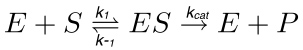 (1)
(1)
حيث ك 1 هو ثابت معدل لتشكيل مجمع ES، ك -1 هو ثابت معدل لتفكك المجمع ES، في حين ك القط هو ثابت معدل الحفاز أو عدد دوران.
وفقا للمعادلة ميكايليس-مينتن 10، ومعدل التفاعل يمكن أن تحسب على النحو التالي:
 (2)
(2)
التي K M = (ك + ك -1 القط) / ك 1 ك والقط = V ماكس / [E]، مع V ماكس كونها السرعة القصوى عندما وصلت كل انزيم منضما إلى الركيزة.
المعايرة المسعر متساوي هو الأداة المستخدمة في هذه الدراسة لوصف الإنزيمية من اليوريا. يرصد هذا الصك من درع ثابت الحرارة التي تحتوي على اثنين صاغ شكل خلايا (الشكل 1). وترتبط هذه إلى الخارج مع أنابيب الوصول الضيقة. يتم تحميل خلية العينة (حوالي 1.4 مل) مع الحل الانزيم، في حين يتم تعبئة الخلية المرجعية عموما مع الماء أو مع المذيبات المستخدمة في التحليل. A حقنة بالتناوب مع إبرة طويلة ومجداف ضجة المرفقة، التي تحتوي عادة كاليفورنيا. 0.3 مل من محلول الركيزة، هي التي شنت على الخلية العينة. يقيس جهاز الحرارية الفرق في درجة الحرارة بين العينة والخلية المرجعية و، وذلك باستخدام "شبكة ردود الفعل الخلية"، فإنه يحافظ على هذا الفارق في صفر عن طريق إضافة أو طرح الحرارة. أثناء التجربة، يتم حقن الركيزة في حل الانزيم عند درجة حرارة اختار ثابتة. عندما ENيأخذ رد فعل zymatic مكان، وكمية الحرارة المنطلقة أو الممتصة يتناسب مع عدد جزيئات الركيزة التي يتم تحويلها إلى جزيئات المنتج. بالإضافة إلى ذلك، فإن معدل تدفق الحرارة ويرتبط مباشرة إلى معدل التفاعل. البيانات المقاسة، والتي تظهر على أنها انحراف من أثر الحرارة من خط الأساس الأولي (الشكل 1)، وتمثل الطاقة الحرارية (μcal / ثانية) التي قدمتها الصك إلى الخلية عينة، والتي تتناسب مع تدفق الحرارة التي تحدث في الخلية عينة مع مرور الوقت.

الشكل 1. التمثيل التخطيطي من المسعر المعايرة متساوي لدراسة التفاعلات الإنزيمية. رد الفعل الأنزيمية التي تحدث عند المعايرة الركيزة (في حقن حقنة) في حل انزيم (في الخلية العينة) النتائج في تغيير ذرقوة القانون النموذجي للتحكيم صادر عن مسعر، اللازمة للحفاظ على الفرق في درجة الحرارة بين الخلية عينة وثابت مرجع الخلية. اضغط هنا لمشاهدة صورة أكبر.
وعموما، فإن تغير الحرارة (س) يتناسب مع المحتوى الحراري المولي للتفاعل (ΔH) وعدد الشامات من الناتج لدت (ن)، والتي بدورها تعطى من قبل مجموع مرات حجم تركيز:
 (3)
(3)
تشكيل المنتج مع مرور الوقت (DP / دينارا)، والتي تتطابق مع معدل التفاعل، وبالتالي يمكن أن تكون ذات صلة لكمية من الحرارة المتولدة مقارنة بنفس الفترة من (DQ / ت) من خلال العلاقة:
 (4)
(4)
وفقا لهذه المعادلة، من أجل الحصول على ميكايليس-منتن مؤامرة فمن الضروري لقياس ط) مجموع المحتوى الحراري ΔH الرحى، والثاني) الحرارة تدفق DQ / دينارا بتركيزات الركيزة مختلفة. عادة، هذه تتم في تجربتين مختلفتين: في التجربة الأولى (الطريقة 1، M1)، يتم حقن الركيزة في حل الانزيم ويتم قياس الحرارة لاستكمال تحويل الركيزة؛ في التجربة الثانية (الطريقة 2، M2)، يتم تنفيذ الحقن متعددة من الركيزة ويقاس معدل إنتاج الحرارة في تركيزات الركيزة مختلفة. هاتين المجموعتين من المعطيات كافية لاستخلاص المعلمات الحركية K M وك القط.
في هذه المادة، يتم وصف بروتوكول العامة لتحديد معالم الحركية للتفاعلات الأنزيمية تنفيذها باستخدام مركز التجارة الدولية. طبقنا طريقة لاليوريا التحلل بواسطة Canavalia الخنجري اليورياحد ذاتها، ونظام مرجعي. اتفاق جيد بين النتائج التي تم الحصول عليها باستخدام هذه الطريقة والبيانات المبلغ عنها في الأدب يدل على موثوقية هذا النهج.
