The combination of the linear sucrose gradient and the ultracentrifugation allows the separation of the different components of the retinal suspension by density. Heavy larger retinal debris and RPE cells sink to or near the bottom of the gradient (Figure 2A). Lighter POS and lighter individual cells or cell debris from the retina migrate as separate bands to reach the top half of the gradient by the end of the centrifugation period. By keeping the eyes and the samples in the dark until after the centrifugation step, the band containing POS can be immediately recognized by its bright orange color. The width and intensity of this band depends on gradient quality and tube height (89 mm height is preferred). Typically, when isolated properly POS should appear elongated, straight or bent when observed on a microscope slide (Figure 2B). Depending on the focus plane on the microscope, they can appear either dark or shiny. If not resuspended properly, they will stay in clumps. If they are damaged (see paragraph below), they will appear as much smaller pieces, a little like a ‘dusty’ background.
Problems that can arise are mostly related to three issues. The first possible issue is that the shaking step of the retinal homogenates is not vigorous enough, which leads to POS remaining attached to PRs thus to decreased yield. The second possible issue is linked to poor gradient casting or gradient quality loss to insufficient stabilization time before loading or excessive tube shaking at any step before the collection. In this case, the bands cannot be properly distinguished and POS cannot be collected in the corresponding tube. Finally, the third potential issue arises if the pH of any of the solutions is wrong: POS membranes may disintegrate and only small POS debris appears on the microscope when visualizing them for yield assessment.
Purified POS can be used for a number of different applications related to studying the phagocytic function of RPE cells. RPE cells are usually used as cell lines or in primary culture from animal models 32,6,12,18-21. More recently, RPE cells derived from reprogrammed stem cells like iPSC or ESC have been used29,30,27,34. Isolated POS serve to test directly the phagocytic ability of mutant RPE cells; e.g., POS are readily phagocytosed by primary RPE cells from wild-type mice, whereas cells from beta5 integrin and MFG-E8 knockout mice phagocytose less POS in the same amount of time6,12. Quantification of POS phagocytosis can be done either by counting FITC-labeled POS manually on microscope pictures6 (Figure 3A), using equipment capable of reading fluorescence intensity10,24,15,35 or a flow cytometer after cell trypsinization36,27. More recently, POS intake and degradation has been evaluated on immunoblots probed for opsins37,38,27.
In vivo, the POS uptake process occurs sequentially as recognition/binding precedes synchronized internalization6. The different phases of uptake can also distinguished in vitro by using appropriate experimental procedures39. Pulse-chase experiments can be performed using temperature switch: POS binding occurs at 20 °C while internalization needs a temperature of 37 °C to proceed32,40,7. This distinction also exists in macrophages, which can recognize POS as well and in which the molecular machinery to eliminate apoptotic cells is very similar to the RPE's machinery7. After FITC-POS phagocytosis, binding and internalization can be quantified in separate samples by quenching the FITC-fluorescence of surface-bound POS during pre-incubation with trypan blue before fixing the cells10. In trypan blue-treated cells, only internal POS will be visualized, and bound POS can be quantified by subtracting internal POS from total POS fluorescence counts. When phagocytosis is evaluated on immunoblots obtained using lysates collected after POS challenge, an equivalent treatment with EDTA before lysis will allow the detachment of POS bound at the cell surface for internal versus total POS phagocytosis comparison37,38.
Tremendous advances have been made in the identification of the phagocytic machinery thanks to experiments employing purified POS. Protein recruitment can be assessed by immunofluorescence co-localization assays13-15,19-21,37,41(Figure 3B). Protein recruitment or activation can be validated on immunoblots after immunoprecipitation or by checking for phosphorylation7,10,12-15,17,19,20,27,35,37,41 (Figure 3C). More open-ended studies on overall gene expression changes after different times of POS challenge have also been performed42.
Isolated POS can also be used to study POS elimination by RPE cells. Indeed, during normal aging and/or when POS are not digested properly, POS degradation products accumulate gradually as lipofuscin deposits and can lead to pathologies due to oxidation mechanisms24,27. Hence, understanding the effects of light and related oxidative damage may require POS challenge of RPE cells18,26. Finally, repeated feeding with normal27 or oxidized25,28 POS can be used to induce cumulative effects in RPE cells in culture in order to understand pathological mechanisms that develop in vivo over longer periods of time.
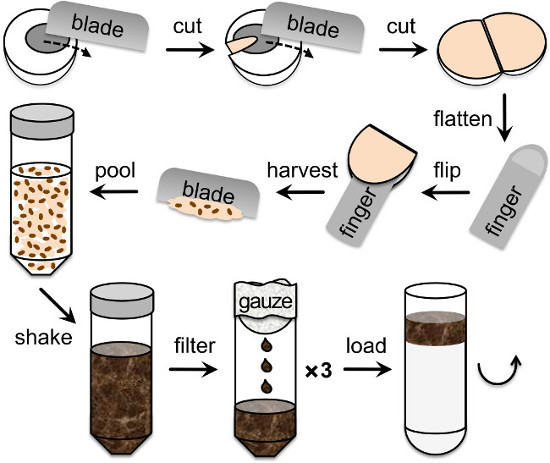
Figure 1. Retina isolation from porcine eyes. Drawing showing the different steps of the eye dissection in order to collect and homogenize the retina before the ultracentrifugation step on continuous sucrose gradients, starting top left. A 60 mm wide blade is used to cut the eye sequentially on one side and then the other in order to be able to stretch it inside-out on a finger tip, thus exposing the retina. Next, the retina is collected by scrapping it from the tapetum with the blade. Pooled retinae are then shaken thoroughly in homogenization buffer, filtered 3 times through double layers of gauze and delicately poured on top of the linear 25 – 60% sucrose gradient.
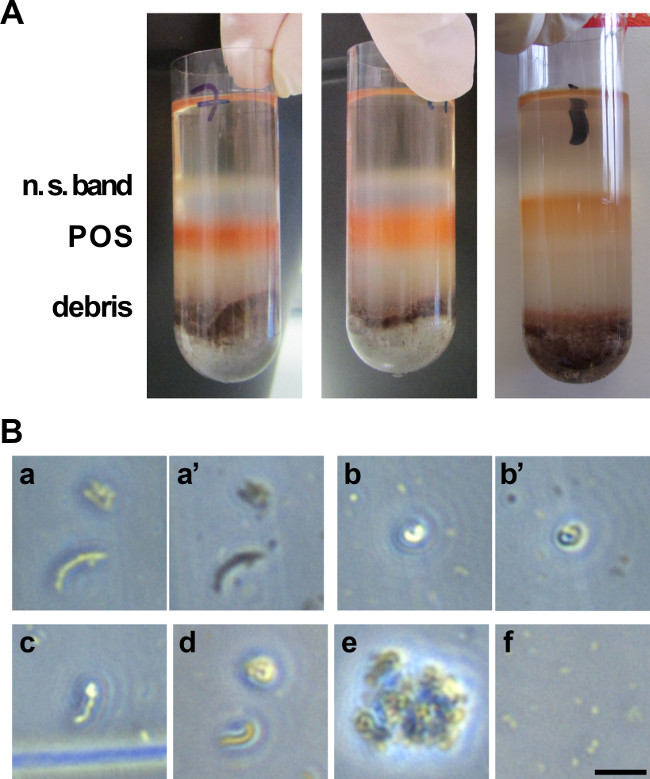
Figure 2. POS purification from porcine eyes. (A) Photograph of the various layers observed on a 25 – 60% sucrose gradient after ultracentrifugation of retina homogenates. The orange band corresponds to POS to be collected. The white upper band corresponds to non-POS related materials, as well as larger debris migrating near or at the bottom of the tube. (B) Picture of isolated POS as observed on a bright field microscope on a counting cell for yield assessment. POS exist in elongated (a, a’, c, d) and whirled (a, a’, b, b’, d) forms. Some elongated POS can display a coiled extremity (c). When changing the plane of focus, POS appearance shifts from shiny to dark (compare a and a’, b and b’). Panel e illustrates POS that have not been resuspended properly and reside in clumps. Panel f shows POS that have been damaged and are not usable, only small pieces are detectable. Scale 10 µm. Please click here to view a larger version of this figure.
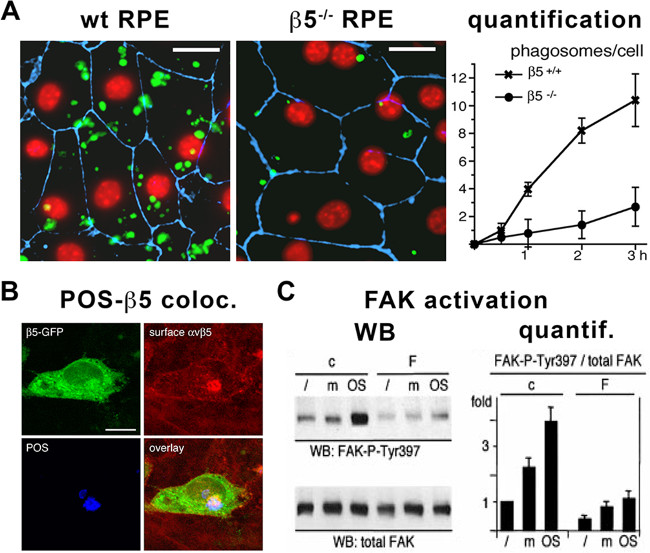
Figure 3. Analysis of RPE phagocytosis using purified POS. (A) Confocal microscope picture and corresponding quantification of the number of phagosomes per cell with time showing that FITC-labeled POS (green) have been less phagocytosed by RPE primary cells isolated from beta5 integrin knockout (β5-/-) compared to wild-type (wt or β5+/+) control mice as indicated. Nuclei (red) were labeled using DAPI and cell junctions (blue) using the tight junction marker ZO-1. Scale bars 10 µm. Reproduced from ©Nandrot et al., 2004, originally published in The Journal of Experimental Medicine 200(12):1539-1545. (B) Confocal microscope pictures showing co-localization (yellow-magenta, bottom right panel) between β5-GFP fusion proteins (green, top left panel), surface αvβ5 integrin receptors (red, top right panel) and POS (blue, bottom left panel). Scale bar 10 µm. Reproduced from Nandrot et al., 2012, originally published in Biology of the Cell 104(6):326-341, ©Portland Press Limited. (C) Immunoblots (WB) and corresponding quantification (quantif.) showing that POS challenge (POS) increases phosphorylation of FAK on the Tyr397 residue when compared without treatment (/) or challenged with medium alone (m) in control cells (c) while cells expressing an inactivated form of FAK (F) do not react, as indicated. Reproduced from Finnemann, 2003, originally published in EMBO Journal 22(16):4143-4154. Please click here to view a larger version of this figure.
| Name of Material/ Equipment | Company | Catalog Number | Comments/Description |
| Specific Material/Equipment | |||
| 2-chamber gradient maker | gradient maker with 30-ml chambers | ||
| 3-mm diameter silicone tubing | tubing for gradient casting | ||
| small size magnetic stir bar | stir bar fitting the gradient maker chamber | ||
| red safelight lamp | inactinic lamp for dissection in the dark | ||
| Ultra Clear 25×89 cm tubes | Beckman | 344058 | ultracentrifugation tubes |
| PP Oak Ridge tubes | Nalgene | 3119-0050 | 30-mL centrifugation tubes |
| Optima LE-80K | Beckman Coulter | 365668 | ultracentrifuge |
| SW 32Ti swing rotor | Beckman Coulter | 369694 | swing rotor for ultracentrifuge |
| Avanti J-26 XP | Beckman Coulter | 393124 | centrifuge |
| JA-25.50 rotor | Beckman Coulter | 363058 | rotor for Avanti J-26 XP centrifuge |
| FITC Isomer I | Life Technologies | F-1906 | fluorescent dye |
| Other Material/Equipment | |||
| counting chamber (such as Neubauer or Malassez) | |||
| dark ice buckets with lids | |||
| scales | |||
| magnetic stirrer and upholding pole | |||
| refrigated microcentrifuge | |||
| 37 °C water bath | |||
| -80 °C freezer | |||
| Consumables | |||
| labcoat | Health and safety | ||
| gloves | |||
| sleeve protectors | |||
| googles | |||
| absorbent pads | |||
| biohazard trash bags and bins | |||
| Weck-Prep blades (60-mm/2.25-in wide razor blades) | Dissection | ||
| 15-cm plastic dish | |||
| sterile gauze sheets | |||
| 15- and 50-mL tubes | Common consumables | ||
| microtubes | |||
| aluminum foil |
Table 1: List of materials and equipments required for the POS isolation protocol.
–>| Stock solution | Storage temperature |
| 70% sucrose | 4 °C |
| 100 mM glucose | 4 °C |
| 1 M MgCl2 | RT |
| 0.5 M taurine | -20 °C |
| 200 mM tris acetate pH 7.2 | RT |
| 200 mM sodium phosphate pH 7.2 | RT |
| 0.1 M NaHCO3 pH 8.4 | RT |
| 0.1 M Na2CO3 pH 11.5 | RT |
Table 1. Recipes of stock solutions and associated storage temperatures.
