Selection:
The H+ killing selection is based on the diffusion of the ammonium weak base, as depicted in Figure 1A. The effect of weak bases and acids diffusion on intracellular pH has been pioneered by Walter Boron and collaborators22. The elegant idea to use this phenomenon to produce a lethal acidification for positive genetic selection was then developed by Jacques Pouysségur17. Under such a protocol, the intracellular pH drops to about 5.5 after the rinse step. Cells which do not express a functional NHE at the plasma membrane cannot recover a neutral pH value and show a typical acidified aspect with a flat shape and a granular cytosol (Figure 1B). After repeated selection cycles (about two H+ killing procedures/week), cells that fully and stably survive this acidification (Figure 1C) emerge, at a frequency of about 10-7. This will result in the formation of individual cellular clones that can be either characterized individually or pooled to obtain cellular populations if desired. Standard experiments such as cell surface biotinylation or RNA silencing can be conducted to control that the protein expressed at the plasma membrane is indeed an intracellular NHE such as NHE7 (see for example Milosavljevic et al.18). As well, RT-PCR and subsequent sequencing should be performed to check for eventual mutations in the sequence of the intracellular NHEs expressed in the selected cell line.
Functional Characterization:
Fluorescence Videomicroscopy:
The activity and ion selectivity of the intracellular NHEs expressed at the plasma membrane can be characterized by measuring the changes in intracellular pH induced by these NHEs in various conditions. For this a videomicroscopy set equipped with illumination and acquisition settings to image a ratiometric probe such as BCECF/AM is schematized in Figure 2A. Figure 2B and 2C show the typical variations in fluorescence values directly obtained for excitations at 490 and 450 nm, following a NH4+ loading acidification. Figure 2D shows the pH curve obtained from these fluorescence values following data treatment described in 2.4).
Lithium Uptake:
Together with hydrogen and sodium, lithium comprises part of the first column of the periodic table and has been shown to be an efficient coupling cation for Na+/H+ exchangers. It is possible to take advantage of this information to setup fast kinetics of lithium uptake, in order to measure in details the functional parameters of Na+/H+ exchangers. As this cation is absent from cell cytoplasm, its accumulation upon cytosolic acidification will quantitatively reflect the activity of the plasma membrane NHE. Moreover, nanomolar to picomolar concentrations of lithium can be measured with great accuracy using atomic absorption spectrometry. Thus, this approach is very powerful when combined with the selection of plasma membrane expressed vesicular exchangers. Figure 3 illustrates the measurement protocol described in 3) of the protocols section. Cells, preferably seeded on multiple-well plates (6 to 24) are acidified as previously described. The kinetic starts with the incubation of the cells in medium containing lithium, and the other cations or inhibitors of interest (Figure 3A).
To stop the uptake, cells are then submitted to four rapid rinses in ice-cold PBS. Operating rapidly removes any extracellular lithium while ensuring that no significant lithium efflux occurs (Figure 3B). Lithium concentration in each well is then measured using atomic absorption spectroscopy (Figure 3C), and initial rates of Lithium uptake, which directly yield the activity of the exchanger at steady-state are obtained as slope measurements of the lithium accumulation time course (Figure 3D).
As for enzyme kinetics, dose-response of initial rates can be used to derive Km values for substrates (Figure 4A) of Ki values for inhibitors. An interesting feature of transporters is that different coupling cations will compete for transport. This yields the possibility of measuring the Km for sodium by competition with lithium uptake (Figure 4B).
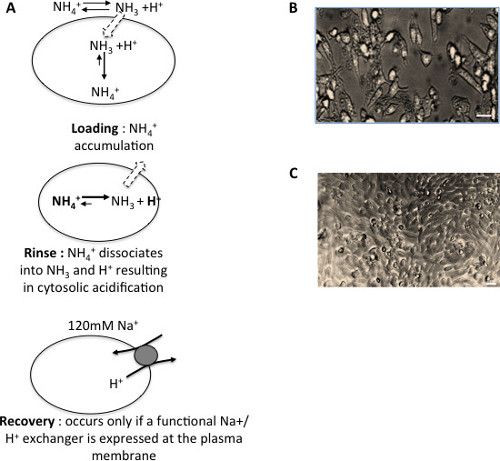
Figure 1: Selection of cells expressing an intracellular Na+/H+ exchanger at their plasma membrane. (A) Strategy for the three steps H+ killing selection of cells expressing a functional non-mutated and non-tagged intracellular NHE at the plasma membrane. (B) Phase contrast microscopy image of PS120 parental cells submitted to an acid load. Scale bar: 25 µm (C) Phase contrast microscopy image of PS120 cells selected for the expression of a vesicular exchanger at the plasma membrane, following an acid load. Scale bar: 25 µm Please click here to view a larger version of this figure.
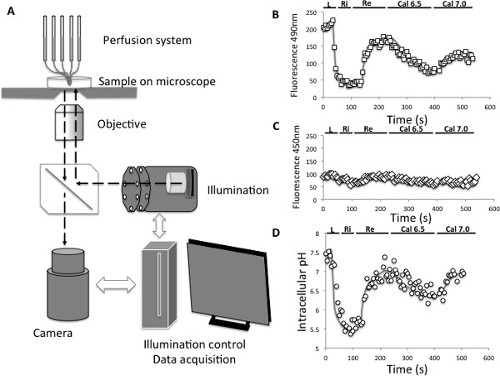
Figure 2: Intracellular pH measurements. (A) Principle of pH videomicroscopy measurement. (B) Evolution of the emitted fluorescence (595 nm) of the BCECF pH probe following an excitation at 490 nm. L: acid load, Ri; Rinse, Re: recovery, Cal6.5: calibration at an intracellular pH of 6.5, Cal7.0: Calibration at an intracellular pH of 7.0 (C) Evolution of the emitted fluorescence (595 nm) of the BCECF pH probe following an excitation at 450 nm. L: acid load, Ri; Rinse, Re: recovery, Cal6.5: calibration at an intracellular pH of 6.5, Cal7.0: Calibration at an intracellular pH of 7.0 (D) Evolution of the intracellular pH computed from the 490/450 nm fluorescence ratios of the BCECF pH probe and from the calibration points. L: acid load, Ri; Rinse, Re: recovery, Cal6.5: calibration at an intracellular pH of 6.5, Cal7.0: Calibration at an intracellular pH of 7.0 Please click here to view a larger version of this figure.
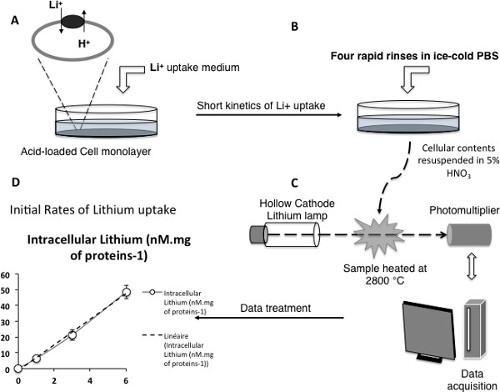
Figure 3: NHE activity measured by initial rates of Li+ uptake. (A–C) Scheme depicting the different critical steps of the measurement technique (D) Example of a measured time course of lithium uptake for the vesicular NHE7 exchanger expressed at the plasma membrane. Note the linearity of the kinetic within the experimental conditions, ensuring initial rates measurement. Please click here to view a larger version of this figure.
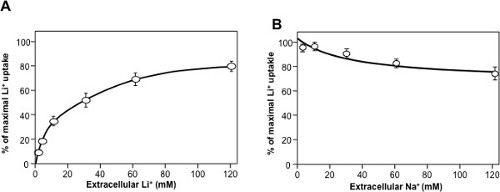
Figure 4: Typical Dose-response curves for NHE7 (original data from18). Initial rates of Lithium uptake were measured using 1-minute kinetics. (A) Dose-response curve for extracellular lithium. (B) Competition of different concentrations of extracellular sodium on 1mM extracellular lithium uptake. Please click here to view a larger version of this figure.
