Figure 1 illustrates the chironomid life cycle; immature stages (egg, larva, pupa) typically take place in, or closely associated with, an aquatic environment. Upon completion of the larval life stage, the larva constructs a tube-like shelter and attaches itself with silken secretions to the surrounding substrate and pupation occurs. Once the developing adult has matured, the pupa frees itself and swims to the surface of the water where the adult can emerge from the pupal exuviae. The exuviae fills with air, and by virtue of an outer waxy layer of the cuticle, it remains floating on the water surface until bacteria begin to decompose the wax layer.
Water currents or wind concentrate floating pupal exuviae into areas of accumulation, such as where riparian vegetation or fallen trees make contact with the water surface, illustrated in Figure 2A. A larval tray and sieve can be used to collect pupal from these natural accumulation areas and evaluate the emergence of Chironomidae from a broad spectrum of microhabitats, as shown in Figure 2B. For certain applications, it is important to collect samples in a consistent, standardized manner so that comparisons can be made among several sample sites or over time at a given sample site. Ten-minute collection periods have been shown to provide adequate evaluations of chironomid relative abundance3,25. For example, Ferrington, et al.3 examined emergence estimates of the species Chironomus riparius and found that estimates did not vary substantially after 12 pan dips were analyzed. Within a 10-min collection period, many more than 12 dips are typically obtained, thus we feel confident that the majority of abundant species within a sample reach will be detected in this timeframe.3
Once SFPE samples have been collected, picked, and sorted, specimens are slide mounted for genus or species identification and creation of voucher specimens. Labeling the slides with appropriate site, collection, and identification information is recommended, as in Figure 3. Typically, the locality label displays information about the country, state, water body, GPS coordinates, study site ID, collection date, and the name of person that collected the sample. Additionally, this label will have a unique slide number for each slide-mounted specimen. The identification label shows the genus and species (when applicable) identification and name of the person that identified the specimen.
Pupal exuviae need to be correctly dissected and oriented for genus identification and voucher specimen preparation. Figure 4A shows the correct dorsal side up pupal exuviae placement on the slide. During placement onto the slide, specimens may not initially lie dorsal side up because they are cylindrical in shape and often filled with ethanol and air bubbles. Therefore, using forceps or a dissection probe to slightly compress the abdomen into the Euparal towards the slide is suggested. Compression should orient the specimen in dorsal view and expel most of the ethanol and air bubbles. Figure 4B demonstrates the dissection that separates the cephalothorax from the abdomen. During this dissection, it is typical for beginners to tear the abdomen between the first and second abdominal segment. Caution should be placed in maintaining the first abdominal segment with the rest of the abdomen. Figure 4C shows the correct dissection and orientation of the pupal exuviae before positioning of the coverslip. For some specimens, it can be difficult to open the cephalothorax so that the suture edges are on opposite sides and the cephalothorax is oriented in ventral view. Again, a slight dorsoventral compression of the cephalothorax to achieve this placement is recommended.
Collections of SFPE have been successfully used in urban lakes in Minnesota to determine accumulation of species (Figure 5A) and genus richness (Figure 5B) and cumulative species composition along a gradient of mean phosphorus concentration/mean lake depth (Figure 6)23. Based on these results, a proof-of-concept study has been implemented for long-term monitoring of Chironomidae in relation to climate change in sentinel lakes across Minnesota (http://midge.cfans.umn.edu/research/biodiversity/chironomidae-slice-lakes/). Rufer and Ferrington23 determined that four SFPE samples per lake per season recovered the majority of the chironomid community and detected important seasonal variation in urban lakes (Figure 5A, B). In all 16 lakes, April samples contained different taxa than May through September samples. Therefore, in northern-temperate regions, sampling four times per season is recommended, with one sample in April and three samples between May and September. However, for different geographic areas and climates, the sampling regime should be tailored to the region to maximize the portion of the community collected.
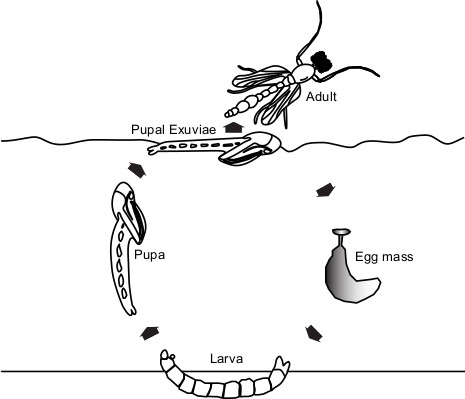
Figure 1. Chironomid life cycle. There are four life stages, egg, larva, pupa, and adult, in the chironomid life cycle. Female adults lay eggs on the surface of the water. Eggs sink to the bottom and typically hatch in several days to one week. After leaving the egg mass, larvae burrow into the mud or construct small tubes in which they live, feed, and develop. Larvae transform into pupae while still in their tubes. After pupation, pupae actively swim to the surface of the water and adults emerge from the pupal exuviae. Please click here to view a larger version of this figure.

Figure 2. Examples of an area of SFPE accumulation and field collection techniques in a stream. (A) An example of where SFPE would accumulate upstream of a log. The white, foamy material is a combination of organic matter, such as macrophytes and algae, and can contain hundreds to thousands of pupal exuviae. (B) An example of how a collector would use a sieve and larval tray to collect SFPE from the riparian banks of the stream. Please click here to view a larger version of this figure.

Figure 3. Diagram showing locations of slide date and locality label (left), identification label (right), and slide mounted pupal exuviae under coverslip (center). Please click here to view a larger version of this figure.
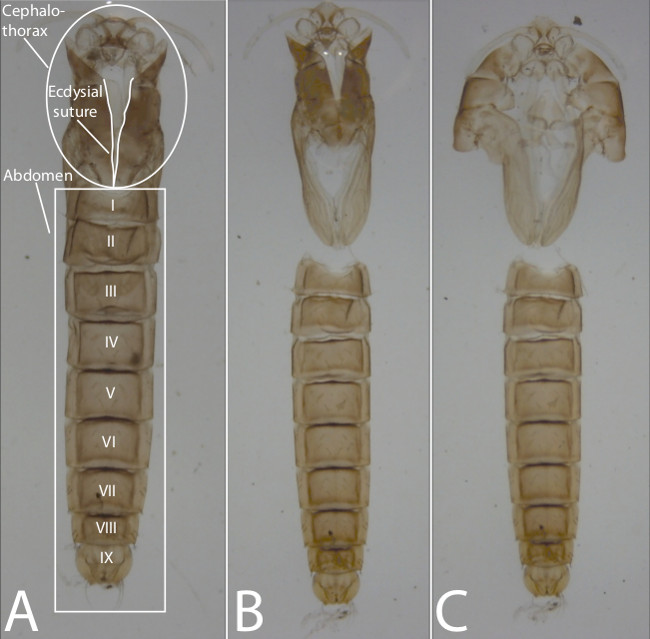
Figure 4. Step-by-step pupal exuviae dissection and orientation. (A) Undissected pupal exuviae (cephalothorax and abdomen with segments numbered in dorsal view). (B) Dissected pupal exuviae (cephalothorax and abdomen in dorsal view). (C) Dissected and oriented pupal exuviae (cephalothorax: ventral view; abdomen: dorsal view). Please click here to view a larger version of this figure.
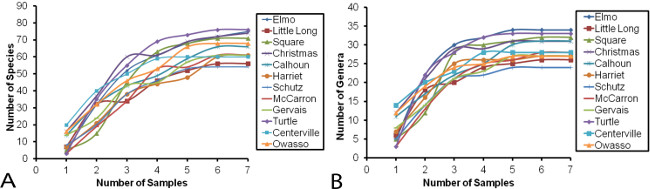
Figure 5: Taxonomic accumulation curves for SFPE samples collected from 16 urban lakes in Minnesota. For both panels, each colored line represents one of the 16 lakes. See Rufer and Ferrington 23 for a detailed description of the characteristics of each lake. Each data point represents a monthly 10-min SFPE sample collected along the downwind shore during the ice-free months of 2005 (April to October). A) Species accumulation curves for SFPE samples. B) Genus accumulation curves for SFPE samples. Please click here to view a larger version of this figure.
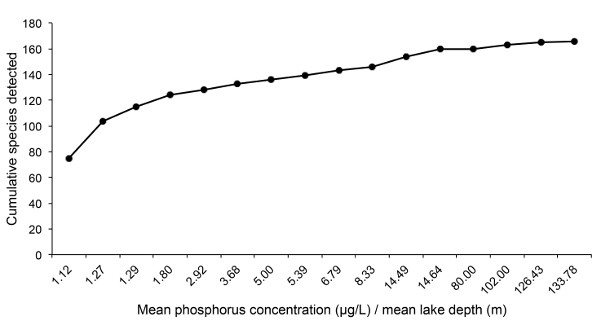
Figure 6: Cumulative species detected across a gradient of lake chemistries from multiple SFPE samples as a function of mean epilimnetic phosphorus concentration (μg/L) over mean lake depth (m) from 16 urban lakes in Minnesota. Each data point represents one of the 16 lakes; lakes are sorted from lowest to highest mean phosphorus/mean depth. See Rufer and Ferrington 23 for a detailed description of the characteristics of each lake. Cumulative number of species encountered increases as the ratio of mean phosphorus concentration over mean lake depth increases.
