الكلوي النقل أيون الظهارية راء iono- العضوي، التنظيم الاسموزي. يقدم ذبابة الفاكهة السوداء البطن مالبيغي (الكلى) أنبوب صغير فرصة لا مثيل لها لدراسة الآليات الجزيئية لنقل أيون الظهارية. ويرجع ذلك إلى مزيج من الوراثة قوية من ذبابة الفاكهة، وإرفاقها مع إمكانية الوصول إلى الأنابيب الكلوية لها لدراسة فسيولوجية هذا. فحص رامزي، الذي سمي على اسم المحقق الذي كان رائدا في تقنية 1، يقيس معدلات إفراز السوائل من الأنابيب مالبيغي معزولة، وتأسست في ذبابة الفاكهة في عام 1994 من قبل شركة داو وزملاؤه 2. هذا مهد الطريق لمزيد من الدراسات باستخدام ذبابة الفاكهة الأدوات الوراثية، مثل نظام GAL4-UAS 3،4، لتحديد مسارات الإشارات خلية محددة تنظم إفراز السوائل. ويشمل على سبيل المثال يشير الكالسيوم في استجابة لهرمون الببتيد 5، وغيرها الكثير 6،7.
وقد أظهرت ve_content "> مزيج من التقنيات الوراثية ودراسة فسيولوجية الكلاسيكية التي الجيل البول في حدوث الطاير من خلال إفراز البوتاسيوم كلوريد الغنية السوائل من الجزء الرئيسي من أنبوب صغير. وهذا يحدث من خلال إفراز بطريق الظهارة مواز من الكاتيونات، في المقام الأول K + ولكن أيضا نا +، من خلال الخلية الأساسية، والكلورين – إفراز من خلال الخلايا النجمية 8-12 القدرة على قياس حدة بطريق الظهارة K + والصوديوم + تدفقات يسمح توصيف أكثر تفصيلا لآليات النقل من قياس إفراز السوائل وحده. على سبيل المثال، في الأنابيب ذبابة الفاكهة unstimulated، نا + / K + إطعام الفئران المانع ابائين ليس له تأثير على إفراز السوائل 2، حتى عندما يتم تثبيط امتصاص حيز الخلايا الرئيسية التي العضوية نقل أنيون المانع توروكولات 13. ومع ذلك، لينتون و أظهر أودونيل أن ابائين depolarizesإمكانات غشاء basolateral، ويزيد نا + تدفق 9. كما هو مبين في ممثل النتائج، ونحن تكرار هذه النتائج، وأظهرت أن K + تدفق هو انخفضت بصورة متزامنة 14؛ زيادة نا + التمويه وانخفض K + تدفق لها آثار معارضة على إفراز السوائل، مما أدى إلى عدم التغير الصافي في إفراز. وبالتالي، هناك قرارين ل"مفارقة ابائين"، أي مراقبة الأولية التي ابائين له أي تأثير على إفراز السائل في أنبوب صغير ذبابة الفاكهة: أولا، في الأنابيب حفز، وتأثير ابائين على إفراز السوائل غير واضح نظرا ل امتصاص من قبل الناقل أنيون العضوية 13؛ وثانيا، في الأنابيب unstimulated، ابائين ومعارضة الآثار على بطريق الظهارة نا + K + والتمويه، مما أدى إلى عدم التغير الصافي في إفراز السائل (انظر ممثل النتائج والمرجع 9). ولذلك، فإن الدور الرئيسي للنا + / K + -ATPبورصة عمان في الأنابيب unstimulated هو خفض داخل الخلايا نا + تركيز لتوليد التدرج تركيز مواتية لنا + -coupled عمليات النقل عبر الغشاء basolateral. في الواقع، عن طريق قياس حدة نا + وK + التدفقات، أثبتنا أن الأنابيب التي تفتقر إلى cotransporter ذبابة الصوديوم والبوتاسيوم-2-كلوريد (NKCC) قد انخفضت K بطريق الظهارة + التمويه، مع عدم وجود مزيد من الانخفاض بعد ابائين بالإضافة إلى ذلك، وأي تغيير في بطريق الظهارة غ + تدفق 14. وأيدت هذه النتائج استنتاجنا أن نا + دخول الخلية من خلال NKCC يتم إعادة تدويرها من خلال نا + / K + إطعام الفئران. وفي مثال آخر، Ianowski وآخرون. لاحظ أن خفض حمام K + تركيز من 10 ملم إلى 6 ملم انخفض بطريق الظهارة K + تدفق وزيادة بطريق الظهارة نا + تدفق في الأنابيب من الرادنة prolixus، مع عدم وجود صافي التغير في إفراز السوائل <sup> 15. كما لوحظت آثار تفاضلية على نا + التمويه وK + تدفق عبر الأنابيب اليرقات في الأنابيب ذبابة الفاكهة ردا على متفاوتة حمية الملح 16 و في نوعين من أنواع البعوض ردا على تربية ملوحة 17.التحدي الأكبر في قياس تدفق بطريق الظهارة أيون في إعداد مقايسة رامزي هو تحديد تركيزات الأيونات داخل السائل المفرز. وقد اجتمع هذا التحدي مع حلول مختلفة، بما في ذلك اللهب photometery 18، واستخدام أيونات المشعة 19، والإلكترون التحقيق الطول الموجي التحليل الطيفي والتشتت 20. هذه التقنيات تتطلب نقل قطرة السائل يفرز إلى أداة لقياس تركيزات أيون. منذ حجم السوائل التي يفرزها ذبابة الفاكهة أنبوب صغير unstimulated صغيرة، وعادة ~ 0.5 NL / دقيقة، وهذا يشكل تحديا تقنيا وأيضا يقدم الخطأ إذا كان بعض من السائل يفرز هوفقدت عند نقله. في المقابل، فإن استخدام أقطاب أيون محددة يسمح بقياس نشاط أيون (من الذي يمكن حساب تركيز أيون) في الموقع. تم تعديل البروتوكول الحالي من تلك المستخدمة من قبل Maddrell والزملاء لقياس بطريق الظهارة K + تدفق عبر أنبوب صغير الرادنة باستخدام valinomycin باسم K + حامل الأيون 21، ويصف أيضا استخدام -butylcalix 4- ثالثي [4]، دآرين tetraacetic حمض رابع إيثيل نا القائم على استر + -specific القطب أيون محددة تتميز Messerli وآخرون. آل 22. كما تم استخدام أقطاب أيون محددة لقياس تركيزات الأيونات في السوائل التي يفرزها الأنابيب مالبيغي في فحص رامزي في الكبار 9،23 واليرقات 16 ذبابة الفاكهة السوداء البطن، ونيوزيلندا جبال الألب يتا (Hemideina الماوري) 24 و 17 في البعوض.
هنا، نحن تصف بالتفصيل استخدام رامزي كماأقول لقياس معدلات إفراز السوائل في الأنابيب مالبيغي من ذبابة الفاكهة، فضلا عن استخدام أقطاب أيون محددة لتحديد تركيزات K + و + نا داخل السائل المفرز وبالتالي احتساب التدفقات بطريق الظهارة أيون. وتقدم لمحة عامة عن الفحص في الشكل 1.
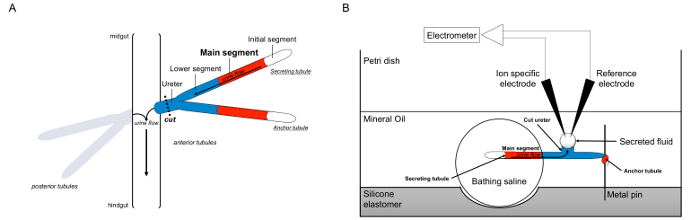
الشكل 1. تخطيطي لمالبيغي الأنابيب الصغيرة والفحص رامزي مع استخدام ايون محددة أقطاب لقياس تركيزات الأيونات. ويوضح هذا الرقم الإعداد لفحص رامزي. (A) كل ذبابة أربعة الأنابيب، وزوج من الأنابيب الأمامية وزوج من الأنابيب الخلفية، التي تطفو في تجويف البطن وتحيط بها الدملمف. في كل زوج، والأنابيب اثنين من الانضمام في الحالب، والتي ثم يفرغ البول عند تقاطع المعي المتوسط وhindguت. الأنابيب عمياء العضوية. يتم إنشاء البول عن طريق القطاع الرئيسي إفراز السائل (كما هو موضح باللون الأحمر)، ويتدفق نحو الحالب والخروج إلى الأمعاء. بعد تشريح، وفصل الزوج أنبوب صغير من الأمعاء عن طريق transecting الحالب. (B) ثم يتم نقل زوج من الأنابيب إلى قطرة من الاستحمام المالحة داخل بئر للطبق الفحص. واحدة من الأنابيب اثنين، المشار إليها هنا باسم "مرساة أنبوبية،" هو التفاف حول مسمار معدني وغير خامل. نبيب الآخر هو أنبوب صغير إفراز. يبقى الجزء الأول (الذي لا تفرز السائل) والجزء الرئيسي من أنبوب صغير إفراز داخل قطرة من الاستحمام المالحة. الأيونات وتحرك المياه المالحة من الاستحمام وداخل تجويف أنبوب صغير من الجزء الرئيسي، ومن ثم تتحرك نحو الحالب، كما يمكن أن تحدث في الجسم الحي. الجزء السفلي (الأزرق) هو خارج المالحة الاستحمام، وبالتالي خاملة. منذ يتم قطع الحالب، السائل يفرز كما يظهر قطرات من نهاية قطع من الحالب. تانه يفرز السائل قطرات يوسع على مر الزمن مع استمرار إفراز، ويتم قياس قطرها باستخدام ميكرومتر بصري. هناك طبقة من الزيت المعدني يمنع تبخر السوائل تفرز. الأقطاب محددة المرجعية وأيون قياس تركيز أيون السائل المفرز. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
