Nierenepithelzellen Ionentransport zugrunde organismal iono- und Osmoregulation. Die Drosophila melanogaster Malpighischen (Nieren) Tubulus bietet eine einmalige Gelegenheit, um die molekularen Mechanismen der epithelialen Ionentransport zu studieren. Dies ist aufgrund der Kombination der leistungsfähigen Genetik von Drosophila, gepaart mit der Zugänglichkeit der Nierentubuli auf physiologische Studie. Die Ramsay Assay, nach dem Forscher, der die Technik 1 Pionier benannt, misst Flüssigkeitssekretion Raten von isolierten Malpighischen, und wurde in Drosophila 1994 von Dow und Kollegen 2 etabliert. Dies ebnete den Weg für weitere Studien unter Verwendung von Drosophila genetische Werkzeuge, wie beispielsweise die GAL4-UAS-System 3,4, um zellspezifische Signalwege regulieren Flüssigkeitssekretion definieren. Ein Beispiel beinhaltet Calcium-Signal in Reaktion auf ein Peptidhormon, 5, unter vielen anderen 6,7.
ve_content "> eine Kombination von genetischen Techniken und klassischen physiologischen Studie hat gezeigt, dass Urin-Generation gezeigt, in die Fliege durch die Sekretion eines Kaliumchlorid-reiche Flüssigkeit aus dem Hauptsegment der Röhrchen erfolgt. erfolgt dies durch die parallele transepithelialen Sekretion von Kationen, in erster Linie K +, sondern auch Na +, durch die Hauptzelle und Cl -. Sekrets durch die Sternzellen 8-12 Die Fähigkeit, transepithelialen K + und Na + Flüsse getrennt zu messen ermöglicht eine genauere Charakterisierung von Transportmechanismen als die Messung Flüssigkeitssekretion allein, z. B. in unstimulierten Drosophila Tubuli, die Na + / K + -ATPase-Inhibitors Ouabain hat keine Auswirkung auf Flüssigkeitssekretion 2, auch wenn ihre Aufnahme in den Hauptzellen wird durch den organischen Anionentransporter Inhibitor Taurocholat 13 gesperrt. Jedoch Linton und O'Donnell zeigte, dass Ouabain depolarisiertder basolateralen Membranpotential und erhöht Na + Fluss 9. Wie in den Repräsentative Ergebnisse gezeigt, repliziert haben wir die Ergebnisse und zeigte, daß K + Flußmittel gleichzeitig um 14; die erhöhte Na + Fluss und verminderte K + Fluss haben entgegengesetzte Auswirkungen auf Flüssigkeitssekretion, was zu keiner Nettoänderung der Sekretion. So gibt es zwei Entschließungen zum "Ouabain Paradox", das heißt, die anfängliche Beobachtung, dass keine Auswirkung auf Flüssigkeitssekretion im Drosophila-Röhrchen Ouabain:. Zunächst in stimulierten Tubuli, ist nicht ersichtlich, die Wirkung von Ouabain auf Flüssigkeitssekretion aufgrund seine Aufnahme durch den organischen Anionentransporter 13; und zweitens, in unstimulierten Tubuli, Ouabain hat gegenläufige Effekte auf transepithelialen Na + und K + Fluss, was zu keiner Nettoänderung der Flüssigkeitssekretion (siehe Repräsentative Ergebnisse und ref. 9). Die primäre Rolle der Na + / K + -ATPase in unstimulierten Tubuli ist es, intrazellulären Na + -Konzentration zu senken, um eine günstige Konzentrationsgradienten zu erzeugen für die Na + -gekoppelten Transportprozesse in der basolateralen Membran. Tat kann durch separates Messen Na + und K + Flüsse zeigten wir, dass Tubuli fehlt die Fliege Natrium-Kalium-2-Chlorid-Cotransporter (NKCC) haben transepithelialen K + Flusses vermindert, wobei keine weitere Abnahme nach Ouabain hinaus und keine Veränderung des transepithelialen Na + Fluss 14. Diese Ergebnisse unterstützten unsere Schlussfolgerung, dass Na + in die Zelle durch die NKCC wird durch die Na + / K + -ATPase recycelt. In einem anderen Beispiel Ianowski et al. Beobachtet, dass die Senkung Bad K + -Konzentration von 10 mM bis 6 mM verringert transepithelialen K + Flußmittel und erhöht den transepithelialen Na + Flusses in Tubuli aus Rhodnius prolixus, ohne Nettoänderung in Flüssigkeitssekretion <sup> 15. Unterschiedliche Auswirkungen auf Na + und K + Flussflusses über Larven Tubuli wurden auch in Drosophila Tubuli als Reaktion auf variierende Salzdiäten 16 und in zwei Mückenarten in Reaktion auf die Aufzucht Salinität 17 beobachtet worden.Die größte Herausforderung bei der Messung des transepithelialen Ionenfluß im Ramsay Probenaufbereitung ist die Bestimmung von Ionenkonzentrationen in der sekretierten Flüssigkeit. Diese Herausforderung hat mit unterschiedlichen Lösungen, einschließlich flamm photometery 18, Verwendung von radioaktiven Ionen 19 und Elektronensonde Wellenlänge Spektroskopie 20 erfüllt sind. Diese Techniken erfordern Übertragung des sekretierten Fluidtropfen auf ein Instrument zur Messung von Ionenkonzentrationen. Da das Volumen der Flüssigkeit durch den unstimulierten Drosophila Tubulus sekretiert ist klein, typischerweise ~ 0,5 nl / min, stellt dies eine technische Herausforderung dar und führt auch Fehler, wenn einige der sekretierten Flüssigkeitverlor bei der Übertragung. Im Gegensatz dazu die Verwendung von ionenspezifischen Elektroden ermöglicht die Messung der Ionenaktivität (von dem Ionenkonzentration berechnet werden kann) in situ. Das aktuelle Protokoll wurde von dem von Maddrellschem und Kollegen zur transepithelialen K + Flusses über die Rhodnius Tubulus Messung mit Valinomycin als Ionophor K + 21 angepasst, und beschreibt auch die Verwendung eines 4-tert-Butylcalix [4] aren-tetraessigsäure Tetraethylester basierten Na + -spezifische ionenspezifischen Elektrode, gekennzeichnet durch Messerli et. al. 22. Ionenspezifischen Elektroden wurden ebenfalls verwendet, um die Ionenkonzentrationen in Flüssigkeit durch Malpighischen im Ramsay Assay bei Erwachsenen 9,23 und 16 Larven Drosophila melanogaster sezerniert zu messen, der Neuseeland-Alpine Weta (Hemideina maori) 24 und in Moskitos 17.
Hier beschreiben wir im Detail die Verwendung des so Ramsaysage Flüssigkeitssekretion Raten in Malpighischen von Drosophila melanogaster, sowie die Verwendung von ionenspezifischen Elektroden, die Konzentrationen von K + und Na + innerhalb von sekretierten Flüssigkeit zu bestimmen und somit die Berechnung der transepithelialen Ionenflüsse zu messen. Eine Übersicht des Tests ist in Figur 1 vorgesehen.
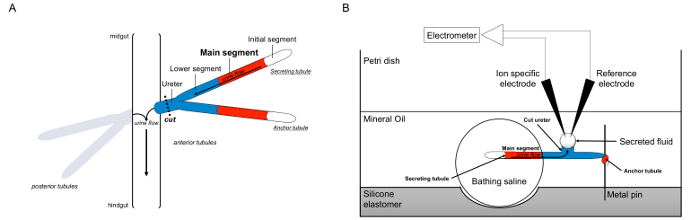
Abbildung 1. Schematische Darstellung des Vasa Malpighi und der Ramsay-Assay unter Verwendung von ionenspezifischen Elektroden, die an Ionenkonzentrationen zu messen. Diese Abbildung zeigt das Setup für die Ramsay Assay. (A) Jede Fliege hat vier Röhrchen, ein Paar von vorderen Tubuli und ein Paar von hinteren Tubuli, die schwimmen in der Bauchhöhle von Hämolymphe umgeben. In jedem Paar, verbinden die beiden Röhrchen in den Harnleiter, die dann leert den Urin an der Kreuzung des Mitteldarms und hindgut. Die Röhrchen sind blind digen. Urin wird durch die (in rot dargestellt) Flüssigkeit absondernden Hauptsegment erzeugt und strömt in Richtung des Harnleiters und hinaus in den Darm. Nach der Präparation wird die Tubuli Paar aus dem Darm durch Durchtrennung des Harnleiters dissoziiert. (B) Das Paar von Röhrchen wird dann in einen Tröpfchenbadesalzlösung innerhalb einer Vertiefung des Assays Schale überführt. Eines der beiden Röhrchen, hier als die "Anker Tubulus" ist um einen Metallstift gewickelt und ist inert. Die andere Röhrchen ist die sezernierenden Röhrchen. Das Anfangssegment (die nicht Flüssigkeit sezerniert) und Hauptsegment der sezernierenden Röhrchen bleiben innerhalb des Tröpfchens des Badens Kochsalzlösung. Ionen und Wasser Umzug von der Badekochsalzlösung und in die Tubuluslumen des Hauptsegments und dann in Richtung des Harnleiters zu bewegen, wie es in vivo auftreten. Das untere Segment (blau) ist außerhalb der Badesalzlösung und daher inert. Da der Harnleiter geschnitten wird, tritt der sekretierten Flüssigkeit als ein Tröpfchen von dem geschnittenen Ende des Ureters. Ter sezerniert Fluidtröpfchen vergrößert Laufe der Zeit als Sekret setzt, und sein Durchmesser wird unter Verwendung eines okularen Mikrometers gemessen. Eine Schicht von Mineralöl verhindert die Verdunstung des sekretierten Flüssigkeit. Die Referenz- und ionenspezifische Elektroden messen die Ionenkonzentration der Flüssigkeit ausgeschieden. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
