腎上皮イオン輸送は、生物のiono-と浸透圧調節の下にあります。 キイロショウジョウバエマルピーギ(腎臓)細管上皮イオン輸送の分子メカニズムを研究するための比類のない機会を提供しています。これは、生理的な研究への尿細管のアクセスとペアにショウジョウバエの強力な遺伝学の組み合わせに起因するものです。技術1を開拓し 、研究者の名にちなんで名付けラムゼイアッセイは、孤立したマルピーギ細管から流体分泌速度を測定し、ダウと同僚2で 1994年にショウジョウバエに設立されました。これは、流体の分泌を調節する細胞特異的なシグナル伝達経路を定義するには、そのようなGAL4-UASシステム3,4 ショウジョウバエ遺伝的ツールを使用して、さらなる研究のために道を開きました。例は、他の多くの中で6,7-ペプチドホルモン5に対する応答におけるカルシウムシグナル伝達を含みます。
ハエは、尿細管の主なセグメントからの塩化カリウムに富む流体の分泌を介して起こるでve_content ">遺伝子技術と古典的な生理学的研究の組み合わせは、その尿の生成を示している。これは主に、陽イオンの並列経上皮分泌を介して行われKは+だけでなく、 の Na + は 、主細胞、およびClを通して– 。星細胞8-12を通じて分泌別途経上皮の K +およびNa +フラックスを測定する能力は、流体分泌の測定よりも輸送メカニズムのより詳細な特性評価を可能にします単独で、例えば、非刺激ショウジョウバエ細管ではNa + / K + -ATPアーゼ阻害剤ウアバインは、主細胞への取り込みは、有機アニオントランスポーター阻害剤タウロコール酸13によって阻害された場合でも、体液分泌2には影響を与えないが、リントンとオドネルは、ウアバインが脱分極することを示しました側底膜電位、およびNa +フラックス9を上昇させます 。代表的な結果に示すように、我々はこれらの結果をレプリケートし、K +フラックス14を同時に減少させることが示されました。増加の Na + フラックスおよび K +フラックスを減少させた分泌の正味の変化が生じ、流体分泌に対する反対の効果を持っています。 。このように、「ウアバインパラドックス」、すなわちへの2つの解像度があり、 ショウジョウバエの細管内の流体の分泌に影響を与えませんウアバイン初期観察は:まず、刺激細管に、流体分泌に対するウアバインの効果は明らかではないが原因有機アニオントランスポーター13によるその取り込み。そして第二に、非刺激細管に、ウアバインは(代表的な結果とrefを参照してください。9)液分泌の純変化なしで、その結果、上皮の Na + および K +フラックスへの影響に反対しています。 Na + / K + -ATPのため、主な役割非刺激細管にアーゼ基底膜を越えて輸送プロセスを共役型のNa +のための有利な濃度勾配を生成するために、細胞内のNa +濃度を低下させることです。確かに、別々 の Na + および K +フラックスを測定することにより、我々は、フライナトリウム-カリウム-2-塩化共輸送体(NKCC)を欠いている細管をさらにウアバイン添加後減少し、経上皮に変化がないがないと、経上皮K +フラックスが低下していることを実証しましたNa +が14フラックス 。これらの知見は、NKCCを通じてセルに入るの Na + をNa + / K + -ATPアーゼを介してリサイクルされ、私たちの結論を支持しました。別の例では、Ianowski ら 6mmの10 mMのからバスK +濃度の低下は、流体分泌の純変化なしで、経上皮K +フラックスとRhodniusのprolixusから尿細管における増加した上皮の Na + 流入を減少することが観察され<sup> 15。幼虫細管全体での Na + フラックスおよび K +フラックスの差動効果も塩分17を飼育することに応答して変動する塩食16に二蚊の種に応じて、 ショウジョウバエの細管において観察されています。ラムゼイアッセイ調製物における経上皮イオンフラックスの測定における最大の課題は、分泌された流体内のイオン濃度の決意です。この課題は、難photometery 18、放射性イオン19を使用すると、電子プローブ波長分散分光法20を含む様々なソリューションと満たされています。これらの技術は、イオン濃度を測定するための機器に分泌液滴の移動を必要とします。非刺激ショウジョウバエ細管により分泌される流体の体積が小さいため、通常は0.5〜NL /分で、これは技術的な課題を提起し、分泌された流体の一部である場合、エラーが導入され転送時に失わ。対照的に、イオン特異的電極の使用は、 その場での (イオン濃度を算出することが可能な)イオン活性の測定を可能にします。現在のプロトコルは、K +イオノフォア21としてバリノマイシン使用してRhodnius細管全体の経上皮K +フラックスを測定するためにMaddrellらによって使用されるものから適合し、また、4- ターシャの -butylcalix [4]アレーン四酢酸の使用が記載されましたテトラエチルエステル系NaがMesserli らによって特徴づけられる特異的イオン固有の電極を+。アル。22。イオン特定電極はまた、大人の9,23および幼虫16 キイロショウジョウバエでラムゼイアッセイにおいてマルピーギ細管により分泌液中のイオン濃度を測定するために使用されてきた、ニュージーランドアルパインウェタ(Hemideinaマオリ )24と蚊17インチ
ここでは、詳細にラムゼイとしての使用を記載ショウジョウバエからマルピーギ細管流体分泌速度を測定するだけでなく、分泌液、したがって、経上皮イオンフラックスの計算内の K +およびNa +の濃度を決定するために、イオン固有の電極を使用すると言います。アッセイの概要を図1に提供されます。
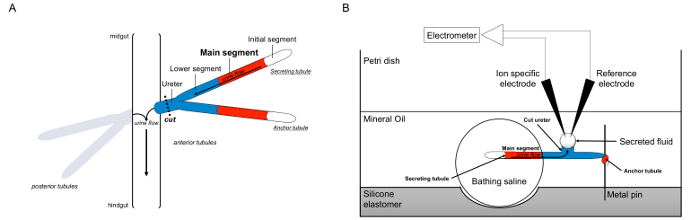
マルピーギ細管の図1の回路図とイオン濃度を測定するためのイオン特定電極の使用とラムゼイアッセイ 。この図は、ラムゼイアッセイのためのセットアップを示しています。 (A)各フライは、4つの細管、前方細管一対の後部細管のペア、血リンパに囲まれた腹腔内のそのフロートを持っています。各ペアでは、二つの細管はその後、中腸とhindguの接合部に尿を空に尿管、に参加しますトン。細管は、ブラインド・エンドです。尿は(赤で示した)は、流体分泌メインセグメントによって生成され、消化管に尿管に向かって内外に流れます。解剖後、細管のペアは、尿管を横断することにより、腸から解離します。 (B)は、尿細管のペアは、その後、アッセイ皿のウェル内入浴食塩水の液滴に移しました。 2細管の一つは、「アンカー細管は、「金属ピンの周りを包み、不活性であるとしてここでいいます。他の細管は、分泌細管です。 (液体を分泌しない)最初のセグメントと分泌細管の主なセグメントは、入浴の生理食塩水の液滴内に残ります。その後、イオンと水の移動入浴生理食塩水から、主要セグメントの尿細管内腔へ、および生体内で起こるように、尿管に向かって移動します。下部セグメント(青)が入浴生理食塩水、したがって不活性外です。尿管が切断されるので、分泌液が尿管の切断端からの液滴として現れます。 T彼は、分泌が続くように、流体液滴が時間とともに拡大し、分泌され、その直径は、接眼マイクロメータを用いて測定されます。ミネラルオイルの層は、分泌液の蒸発を防止します。リファレンスとイオン特定電極が分泌液のイオン濃度を測定する。 この図の拡大版をご覧になるにはこちらをクリックしてください。
