Почечная транспорт эпителиальных ионов лежит в основе организменном ионосферы и осморегуляции. Дрозофилы мальпигиев (почечная) трубочка предлагает уникальную возможность для изучения молекулярных механизмов транспорта ионов эпителия. Это связано с комбинацией мощных генетики дрозофилы, в паре с доступностью его почечных канальцев к физиологическому изучению. Рамсей анализ, названный в честь исследователя, который впервые технику 1, измеряет жидкость цены секреции из изолированных мальпигиевых канальцев, и была создана в 1994 году дрозофилы в Доу и коллег 2. Это проложило путь для дальнейших исследований с использованием генетических Drosophila инструменты, такие как системы GAL4-UAS 3,4, чтобы определить соты сигнальные пути, регулирующие секрецию жидкости. Пример включает в себя сигнализацию кальция в ответ на это пептидный гормон, 5 среди многих других 6,7.
ve_content "> Сочетание генетических методов и классического физиологического исследования показали, что поколение мочи в лету происходит через секреции хлорида богатых жидкости калия из основного сегмента канальцев. Это происходит через трансэпителиального секреции параллельно катионов, прежде всего, К +, но и Na +, через основной камере, и Cl -. секреции через звездчатого клетки 8-12 Способность отдельно измерить трансэпителиальная потоков К + и Na + позволяет более подробную характеристику транспортных механизмов, чем измерение секреции жидкости в покое. Например, в нестимулированных канальцев Drosophila, Nа + / К + -АТФазы ингибитор оуабаин не имеет никакого эффекта на секрецию 2 жидкости, даже когда его поглощение в основных клетках ингибируется органический анион транспортер ингибитора таурохолата 13. Тем не менее, Линтон и О'Доннелл показал, что уабаин деполяризуетбазолатеральный мембранный потенциал, и увеличивает поток Na + 9. Как показано в представительных результатов, мы воспроизвели эти результаты и показали, что поток К + одновременно снизился 14; увеличение потока Na + и K + снизился поток имеет противоположные эффекты на секрецию жидкости, в результате чего не чистого изменения в секреции. Таким образом, есть две резолюции к "уабаина парадокса", т.е., начальное наблюдение, что уабаина не имеет никакого эффекта на секрецию жидкости в канальцах Drosophila:. Во-первых, в стимулированных канальцев, эффект уабаина на секрецию жидкости не является очевидным из-за его поглощение в органический анион транспортер 13; во-вторых, в нестимулированных канальцев, оуабаин имеет противоположные эффекты на трансэпителиальная Na + и К + потока, что приводит к отсутствию изменений в чистой секреции жидкости (см Представитель Результаты и Ref. 9). Таким образом, основная роль Na + / K + -АТФазы в стимулированных канальцев снизить концентрацию внутриклеточного Na + для создания благоприятного градиента концентрации для Na +, -coupled транспортных процессов по базолатеральной мембране. Действительно, отдельно измерения Na + и K + потоки, мы показали, что канальцы не хватает на лету натрия калия-2-хлорида котранспортера (NKCC) снизились трансэпителиальный K + поток, без дальнейшего снижения после уабаина того, и никаких изменений в трансэпителиального Na +, поток 14. Эти данные поддержал наш вывод, что Na +, введя ячейку через NKCC возвращают через Na + / K + -АТФазы. В другом примере, Ianowski и др. Наблюдали, что снижение ванны концентрации К + от 10 мм до 6 мм уменьшилось трансэпителиальный K + поток и увеличение трансэпителиальный Na +, в канальцах поток от Rhodnius prolixus, без чистого изменения в секреции жидкости <sup> 15. Дифференциальные воздействие на поток и K + Na + поток через личиночной канальцев наблюдали также в Drosophila канальцев в ответ на разной соли диеты 16 и в двух видов комаров в ответ на выращивании соленость 17.Самой большой проблемой в измерении потока ионов трансэпителиальная в подготовке Рамсей анализа является определение концентраций ионов внутри секретируется жидкости. Эта задача была выполнена с различной решений, в том числе фотометрию пламени 18, применение радиоактивных ионов 19, и волны зонда электронного дисперсионные спектроскопии 20. Эти методы требуют передачи секретируемого капли жидкости на прибор для измерения концентрации ионов. Поскольку объем жидкости, секретируемый нестимулированных Drosophila трубочку мала, обычно ~ 0,5 нл / мин, это создает техническую проблему, а также вносит ошибку, если некоторые из секретируемого жидкостипотеряли при передаче. В противоположность этому, использование ион-специфических электродов позволяет измерять активности ионов (из которой концентрация ионов может быть рассчитано) на месте. Текущий протокол был адаптирован от используемого Maddrell и коллегами для измерения трансэпителиальный K + поток через трубочку Rhodnius в использовании валиномицина как K + ионофор 21, а также описывает использование 4-трет–butylcalix [4] арена-этилендиаминтетрауксусной кислоты тетраэтилсвинец на основе сложного эфира Na + Определённые ионный конкретных электрод характеризуется Messerli др. др. 22. Ион-специфичные электроды были также использованы для измерения концентрации ионов в жидкости, выделяемой мальпигиевых канальцев в анализе Рамсей во взрослой 9,23 и личинок дрозофилы 16, Новой Зеландии Горные Weta (Hemideina маори) 24 и в 17 комаров.
Здесь мы подробно использование Рамсей какговорят, чтобы измерить жидкости цены секреции в канальцах мальпигиевых из дрозофилы, а также использование ион-специфических электродов для определения концентрации K + и Na + в секретируемого жидкости и, таким образом расчет трансэпителиальная ионных потоков. Обзор анализа обеспечивается на фиг.1.
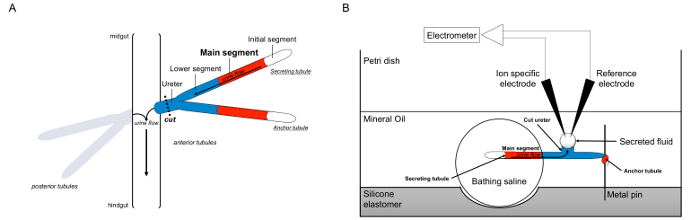
Рисунок 1. Схема мальпигиевых и Рамсей Анализ с использованием ион-специфических электродов измерить концентрации ионов. Эта цифра показывает установку для анализа Рамсей. (А) Каждый муха имеет четыре трубочки, пару передних канальцев и пару задних канальцев, которые плавают в брюшной полости, окруженной гемолимфы. В каждой паре, две трубочки соединяются в мочеточнике, который затем впадает мочи на стыке кишки и hindguт. Трубки слепы состава. Моча образуется в жидкости, секретирующие основной сегмент (показан красным), и течет к мочеточника и вышел в кишечнике. После вскрытия, пара трубочка диссоциирует из кишечника по пересекающих мочеточник. (B) пара канальцев затем переносили в капле физиологического раствора купания в лунку анализа блюдо. Один из двух трубочек, называют здесь как "якоря трубочку," обернут вокруг металлический штырь и инертен. Другой трубочка является секрецию канальцев. Начальный участок (который не секретирует жидкость) и основной сегмент, секретирующих трубочку оставаться в капле физиологического раствора купания. Ионы и вода переход от купания и солевом растворе в просвете канальцев основной сегмент, а затем перейти к мочеточника, как это происходит в естественных условиях. Нижний сегмент (синий) находится за пределами купания физиологического раствора, и поэтому инертны. Поскольку мочеточник вырезать, секретируемый жидкости возникает как капли от срез мочеточника. ТОн секретируется капель жидкости увеличивает течением времени, как секреции продолжается, и его диаметр измеряется с помощью окулярного микрометра. Слой минерального масла предотвращает испарение жидкости секретируемого. Конкретные ссылки и ионов электроды измерения концентрации ионов секретируемого жидкости. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
