En mer grundläggande förståelse för hur T-celler känner igen antigen kräver tittar på rätt ställe, det vill säga inom den immunologiska synapsen bildas mellan T-cellen och APC. Här är molekylära bindande kinetik inte bara bestäms av de inneboende biokemiska egenskaperna hos interaktions inblandade parterna utan beror i stor utsträckning på cellulära parametrar, som omfattar cellulära krafter, membran arkitektur och laterala interaktioner mellan membranproteiner samt synaps specifika geometriska begränsningar 3. Biokemiska metoder är begränsade i upplösningsförmåga som de nödvändiggör avbrott i åtminstone en av de synaptiska membranen inblandade. Av denna anledning en FRET-baserad avbildning metodik utvecklades för att övervaka bindning av TCR till antigena pMHCs 2. Här T-celler är inredda med en rekombinant och platsspecifikt märkt TCRβ-reaktivt enkelkedjigt antikroppsfragment (SCF V) och konfronteras med planen ar glas stöds lipiddubbelskikt (SLBs), som hyser MHC klass Il-molekyler laddade med en fluorescensmärkt antigen peptid, samstimulerande molekyler och vidhäftningsproteiner. Synaptic bindning mellan färgämnesmärkta TCR och färgämnesmärkta pMHC resulterar i FRET, som kan övervakas på en bulk och enda molekyl nivå genom total inre reflektion fluorescens (TIRF) mikroskopi.
I denna artikel förklaras i detalj hur man använder SLBs för analys T-cells synapser, verifiera deras integritet genom en funktionell T-cellskalciumflödesanalys, beteende FRET mätningar i bulk och med enda molekyl känslighet och analysera insamlade data. Rekommendationer erbjuds till att producera riktigt överensstämde proteiner som krävs för dubbelskiktsfunktionalisering. För mer specifik information om dubbelskiktsbildning och inställning av en lämplig TIRF mikroskop hänvisas till en extra allmänhetens tillgång JUPITER publikation publiceras rygg mot rygg 4.
tält "> Naturen av SLBsFunktionaliserbara SLBs kan lätt genereras från unilamellära vesiklar (SUV) innehållande de två lipidema 1-palmitoyl-2-oleoyl-sn-gylcero-3-fosfokolin (kort: POPC, 90-99%) och 1,2-dioleoyl- sn – glycero-3 – {[N- (5-amino-1-karboxipentyl) iminodiättiksyra] -succinyl} (kort: DGS NTA-Ni, 1-10%). SUV sprids på rena glasskivor för att bilda en sammanhängande plan tvåskiktsmembran 4. DGS-NTA-Ni tjänar till att förankra polyhistidin-märkta proteiner via polyhistidin medierad komplexbildning med den syntetiska NTA-Ni-haltiga huvudgruppen (Figur 1A). För stabil association ett ersätter typiskt den nativa transmembrandomänen och den cytoplasmiska svansen hos adhesionsproteinet ICAM-1 och den samstimulerande molekylen B7-1 med en tagg som innehåller tolv histidiner (ICAM-1-12H, B7-1 -12H) (Figur 1B) . Den peptidladdade klass II molekyl IE k innehåller två membraninbäddade (α och β) polypeptidkedjans. De trans / cytoplasmadomäner av båda kedjorna måste ersättas med en etikett som innehåller sex histidiner vardera (IE k α 6H β 6H eller IE k -2x6H). Som ett alternativ, förlängning av α-kedja med tolv histidiner och lämnar den extracellulära domänen av β-kedjan omärkta (ger upphov till IE k α 12H β 0H eller IE k -12H) ger upphov till tillfredsställande resultat (figur 1B).
Platsspecifik märkning av pMHCs
Det är viktigt att märka pMHC stökiometriskt och platsspecifikt för att kunna omvandla uppmätta FRET avkastningen till meningsfulla jämviktsbindningskonstanter. Detta kan uppnås genom kemisk märkning av en syntetisk peptid som laddas in i peptidbindningsklyftan av rekombinant histidin-taggade MHC klass Il-molekyler 2,5. Peptiden inkluderar alla rester av T-cellepitop som viktell som en kort C-terminal linker (GGS) följt av cystein (t.ex. i mal cytokrom c (MCC) peptid ANERADLIAYLKQATK- GGSC, linkern är markerad i fet stil). Denna cystein används för att märka peptiden stökiometriskt med användning av maleimid-dye-derivat. Vid det här laget extra omsorg bör ägnas åt att verifiera kvantitativ färg koppling till cystein-peptid. HPLC-rening av peptiden-dye addukten rekommenderas och måste följas av elektrosprayjonisering masspektroskopi. Alla inspelade massa som motsvarar peptiden edukt (utan färg) återspeglar ofullständig märkning. Om detta är sant, bör HPLC-renad peptid underkastas konsekutiva rundor av färgmärkning tills märkningen bedöms kvantitativt. Notera att MALDI-TOF-masspektroskopi bör undvikas eftersom denna metod innefattar laserstrålning för prov jonisering. Denna behandling sönder bifogade känsliga fluoroforer innan peptidmassa utläses och därmed underrepre senterar grader av färgämne-konjugering.
Indirekt men sätesspecifik märkning av cellbundna TCR med användning av monovalenta enkelkedjiga Fv-fragment
Det är fortfarande en utmaning att fästa färgämnen för att cellyte-associerade proteiner av levande celler i ett ställesspecifikt sätt. För att övervinna detta hinder för ytexponerade TCR har en monovalent enkelkedjigt version (scFv) från generna enligt TCRβ -reactive monoklonala antikroppen H57-197 2 konstruerats. Kristallstrukturen för denna antikropp i komplex med TCR tillåter att rationellt utforma en version, i vilken en serinrest i omedelbar närhet till C-terminalen av MHC associerad peptid (där motsvarande FRET partnern färgämne är fäst) substitueras för en cysteinrest. Denna mutant cystein fungerar sedan som en acceptor för färg konjugering (Figur 2).
Metoder för att spela FRET
nt "> Bulk FRET värden är bäst lämpade för att kontrollera förhållandet mellan valda mellan färg avstånd och FRET effektivitet mäts i denna TCR-pMHC bindande system 2. Dessutom bulkmätningar FRET avslöja kvalitativa och kvantitativa skillnader i synaptiska TCR-pMHC tillhörighet ( se nedan och protokoll avsnitt 3.2). Olika metoder för att kvantifiera FRET effektivitet har införts i litteraturen 6. I den här artikeln FRET registreras via(a) donator återhämtning efter acceptor blekning, och via
(b) strålningskänsliga FRET acceptor utsläpp.
Den första metoden (a) kräver användning av ett FRET-acceptor som lätt kan photobleached, och en givare, som är ganska fotostabila. Dessutom är det viktigt att se till att photobleached acceptorn är inte längre kan släcka donatorfluorescensen. Eftersom samma detekteringskanalen (donator) används för kvantifiering, faktorer ingen korrigering end inga kromatisk aberration måste beaktas, vilket gör denna metod enkel och tillförlitlig. Däremot kan kvantitativa mätningar inte upprepas på samma prov plats och förändringar i FRET kan inte spelas in under tiden. För att undvika effekter orsakade av molekylär diffusion eller cellulär motilitet snabb blekningssteg bör eftersträvas, vilket minimerar tiden passerar mellan första FRET donator (före acceptor blekning) och den andra FRET donator bild förvärv (efter FRET acceptor blekning). Det är rekommendera att använda en kraftfull laser ljuskälla av den FRET-acceptom excitationsvåglängden för att minimera dekoration och blekningstider.
I motsats, i tillvägagångssättet sensibiliserade mätning FRET utsläpp (b) FRET donatorn är glada och utsläpp av FRET mottagaren observeras i FRET acceptkanalen. Förändringar i FRET acceptor signal kan spelas in med tiden, men utsläpp av FRET donatorn i rödskiftade acceptor kanal (termed genomblödning) och FRET acceptor tvär excitation via givarexcite måste noggrant bestämmas och subtraheras från den inspelade FRET acceptor kanal. För detta motsvarande FRET donator och FRET-acceptor-bilder måste spatialt inriktade.
Detektion av en enda molekyl (sm) FRET händelser
Med användning av laser som exciteringskälla, en känslig kamera och buller-försvagade TIRF mikroskopi fluorescensen av enstaka fluoroforer lätt kan spåras över tiden. Liknande gäller för detektion av intermolekylära smFRET händelser. Dock kan komplikationer orsakas av FRET donator genomblödning och korsexcitering av FRET mottagaren, och därmed stor försiktighet måste iakttas vid justering av fluoroforenhetema tätheter i smFRET experimentet.
I protokollet nedan (protokoll avsnitt 4) TCR valdes FRET givare i hög förekomst och pMHC som FRET acceptor i begränsad omfattning. Att dämpa FRET donator genomblödning tillräckligt, dekorera 10-30% av TCR med fluorescerande scFv och 90-70% av TCR med icke-fluorescerande scFv. Här FRET acceptkanalen valdes som enda molekyl kanal eftersom det är konfokal med enda molekyl FRET kanal. Detta bidrar till att rikta smFRET händelser med enda molekyl FRET-acceptorer, som är grunden för smFRET validering.
Utvinna synaptiska off-hastigheter genom smFRET mätningar
Fotoblekning av både FRET givare och FRET acceptor måste redovisas vid extrahering halveringstiden interaktioner från enda molekyl FRET spår. Antalet observerbara FRET-signaler i början av sitt utseende som en enda donator-acceptor par N (0) minskas med tiden genom både ej bindande för receptor-ligandkomplex och fotoblekning. Antalet överlevande komplex vid en given tidpunkt N (t) kan matematiskt uttryckas på följande sätt:
<p class = "jove_content">
I fotoblekning termen exp (-t / τ blekmedel) tiden t beskrivs av produkten av antalet observationer n och belysningstiden t sjuk på grund av den icke-kontinuerliga, diskreta observationsmod (dvs., blekning endast sker under belysning ). Inom den kinetiska termen exp- (t / τ av) tiden t är produkten av antalet observationer n och tiden t fördröjningen för en enstaka FRET iakttagelse (dvs, händer kinetisk ej bindande kontinuerligt). Ekvation 1 kan uttryckas som:
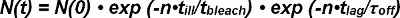
Termen τ blekmedel / τ sjuk describes antalet observationer tills blekning inträffar och definieras som väntevärdet <n bleka> sin exponentiell funktion. Ekvation 2 kan förenklas enligt följande:
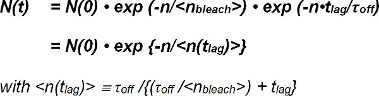
Väntevärdet <n (t eftersläpning)> av antalet bildrutor N (t) med observerbara FRET-händelser efter tiden t direkt bestäms från experimentet. Det beror på den inställbara tiden mellan observationer (t LAG) väljs i försöket och de okända värdena för τ off (inversen av off-rate k off) och <n bleka>, det förväntade värdet av antalet observationer innan blekningen sker .
Således, beräkning av väntevärdet <n (t eftersläpning)> under åtminstone två värden på t <sub> lag tillåter experimentell bestämning av <n bleka> och t bort.
Extrahera synaptiska 2D-K D-värden genom FRET baserade mätningar
Mätning TCR beläggning a, det vill säga förhållandet mellan bundna TCR och totala TCR, är centralt för att avgöra synaptiska 2D-K D-värden. Enligt ekvation 4 denna term är direkt proportionell mot den uppmätta FRET erhållande så länge TCR tjänar som FRET givare och pMHCs som FRET-acceptorer.
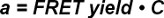
med a = TCR beläggning, C = omvandlingsfaktor
C är en konstant, som beror av FRET-systemet och fluoroforerna som används. Det kan bestämmas experimentellt, såsom visas nedan. en kan omvandlas till en 2D-Kd enligt ekvation 5, närinitial densitet av TCR-ligander före tillsats av T-celler till dubbelskiktet är känd. Detta är på grund av den höga mobiliteten hos SLB lösa proteiner och även eftersom SLBs ger en nästan outtömlig reservoar av ligander 2.
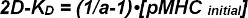
med [pMHC initial] = initial densitet av pMHC före tillsatsen av T-celler
Med ekvationerna 4 och 5 kan man nu enkelt bestämma den synaptiska 2D-K D mellan TCR och pMHC. Detta är mest tillförlitligt sker med FRET mätningar baserade på givar återhämtning efter acceptor blekning (se protokoll avsnitt 3.1).
Men för att mäta C förhållandet mellan FRET-intensiteten I FRET (korrigerat för bakgrunds, FRET-givare genomblödning och FRET acceptor tvär excitation) och TCR beläggning en måste bestämmas. För detta, enbehöver veta förhållandet R mellan den genomsnittliga fluorescensintensiteten av enstaka TCR-associerade FRET givar fluoroforer (t ex Cy3 eller AF555) sm jag FRET givare och den genomsnittliga intensitet enda molekyl FRET händelser sm jag FRET. R beror på FRET-systemet i fråga, emissionsfilter och kamera som används för fluorescensdetektering.
TCR inflyttning en kan då direkt bestämmas enligt ekvation 6.

med R = sm jag FRET donator / sm jag FRET
R bestämdes som 1,45 för H57 scFv- Cy3 / pMHC-Cy5 systemet leder till:
a = bulk jag FRET / bulk jag TCR-Cy3 • 1,45
Förhållandet mellan TCR beläggning en och FRET utbytet kan bestämmas genom FRET donator återvinnay efter acceptor blekning. För detta båda parametrarna är avsatta mot en annan för ett antal TCR microclusters såsom visas i fig 4A .Den Lutningen på den linjära passform anger omvandlingsfaktorn C (ur ekvation 4).
Såsom visas i figur 4A, uppgår för (a) H57 scFv C – Cy3 / pMHC-Cy5 FRET-systemet och (b) den applicerade mikroskopsystemet konfiguration till 1,995. TCR inflyttning en lätt kan härledas på följande sätt:
TCR beläggning a = FRET ge • 1.995
