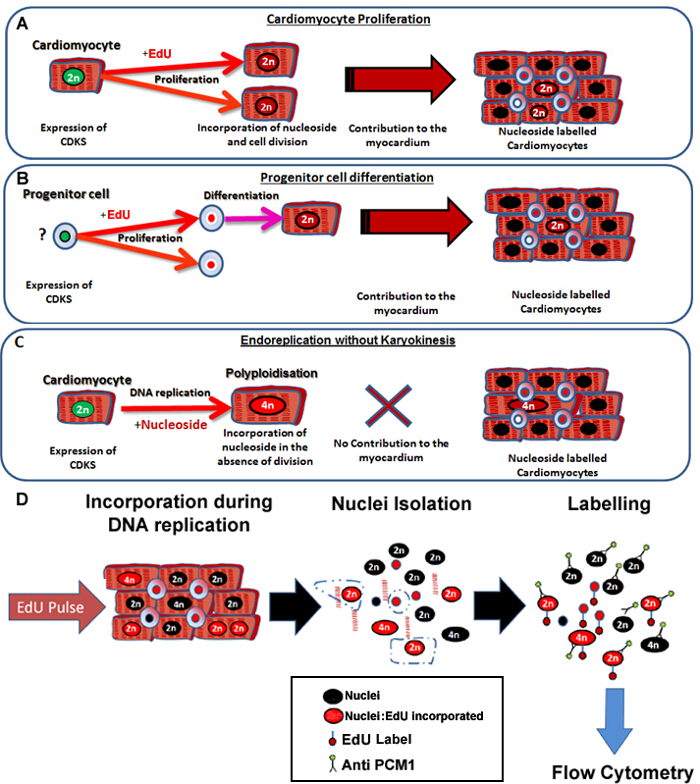
Работа для животных была утверждена и одобрена наблюдательным советом Университета Ньюкасла по этике. Все процедуры на животных были проведены в соответствии с руководящими принципами из Директивы 2010/63 / ЕС Европейского парламента по вопросу о защите животных, используемых для научных целей. 1. Администрация EDU Растворить в ОБР стерильным физиологическим раствором (0,9% вес / об) при конечной концентрации 12,5 мг / мл. Раствор нагревают до 40 ° С и вихрем для того, чтобы полностью раствориться. Хранить достаточное количество ОБР / физиологическом растворе при 4 ° С, чтобы впрыснуть всех мышей в исследовании, что позволяет в течение 6 ежедневных инъекций. Примечание: Как правило более 6 мышей требуются для каждой экспериментальной группы. Однако расчеты мощности должны быть выполнены для независимых исследований. Взвешивание отдельных мышей. Рассчитать необходимый объем для администрирования 100 мкг / г EDU / солевого раствора к каждой мыши (8 мкл акций Edu раствора на грамм). Проведем соответствующий Volumе / Edu физиологический раствор в шприц инсулина с иглой 25 G. Выполните внутрибрюшинно (IP) инъекции при обработке мышей с использованием утвержденных методов домашнего офиса. Поместите мышь на крышке клетки. Осторожно потяните на основание хвоста и крепко возьмитесь мышь за шкирку. Выставляют живота для инъекций, держа мышь с указательным и большим пальцем у основания головы и наклоняя голову животного назад по направлению к полу. Протрите живота с 70% -ным раствором спирта. Найдите среднюю линию животного и нижний правый квадрант. Вводят раствор EDU / физиологического раствора животного в нижний правый квадрант. Вводят мышей с EDU / солевом растворе и фиксируют время суток. Повторите шаги от 1,2 до 1,4, в то же время суток, в течение 6 дней подряд. Примечание: В качестве отрицательного контроля Эду требуется установить стробирования для анализа проточной цитометрии, вводят дополнительный возраст, пол и экспериментально совпадаютред мышь с эквивалентным объемом физиологического раствора без Эду. Это обеспечит необходимый контроль (без управления EDU). Этот контроль должен быть включен для каждого исследования, чтобы учесть различия в партиях реагентов и цитометра настройки. 2. Сбор сердца Через 24 ч после 6 – й инъекции Edu, пожертвуйте мышей с помощью цервикальной дислокации (или альтернативного графика , утвержденного 1 Home Office метод). Поместите жертвенного животного в положении лежа на спине и сделать разрез кожи от середины живота к диафрагме с хирургическими ножницами. Вырезать диафрагму, и удерживая грудину от полости тела, срезанные на двусторонней основе, чтобы обнажить сердце. Слегка приподнимите сердце и перерезать основные кровеносные сосуды тракта оттока рассекать сердце из грудной полости. Сразу же место каждое сердце в физическому лицу 15 мл центрифужную пробирку, содержащую 10 мл PBS, охлаждали до 4 ° С. Передача сердца SEPлярном 10 см чашки Петри, используя скальпель вырезать каждое сердце в двух в сагиттальной ориентации и моют сердце, осторожно сжимая с парой щипцов. Затем замените PBS, чтобы удалить как можно больше крови, насколько это возможно. Повторите этот шаг дважды. Поместите обе части каждого сердца в той же индивидуальной 1,5 мл микроцентрифужных трубки. Переходите к шагу 3 или альтернативно хранить образцы -80 ° C, как описано в пунктах 2.9 и 2.10. С помощью 25 G иглы, сделать небольшое отверстие в крышке микроцентрифужных трубки, чтобы предотвратить крышку от отверстия за счет расширения газов, когда сердце обрабатывается. Добавьте каждый микроцентрифужных трубки и поместить в емкость с жидким азотом для замораживания. 3. Ядра диссоциация и кардиомиоцитов Ядра Этикетировочное Примечание: Это ядра маркировки диссоциации и кардиомиоцитов ядер приспособлена от такового Бергманн и др 11 Выполните эту процедуру на всех образцах инжектированных.с Эду и солевым раствором закачиваемой (отсутствие управления EDU) отрицательного контроля. Приведены в таблице 1 для рецепта решений , необходимых для выполнения этих шагов. Для каждого сердца, чтобы проанализировать, поместите 36 мл 1% BSA / PBS раствора в ультрацентрифужную пробирку, инкубировать в течение 30 мин, вылить, а затем дайте высохнуть на воздухе. Примечание: Эта трубка используется на этапе 3.6. Поместите отдельные замерзшие сердца на 10 мм блюдо на льду и пропустить через мясорубку, используя скальпель, позволяющий сердца размораживать в процессе мясорубки. Поместите фарш сердца в 15 мл буфера для лизиса клеток в центрифужные пробирки объемом 50 мл и гомогенизируют с помощью гомогенизатора зонда в течение 15 секунд при 25000 оборотов в минуту при комнатной температуре. Добавить еще 15 мл буфера для лизиса клеток в пробирку центрифуги и переносят в 40 мл гомогенизатора Даунса с большим зазором пестиком. Выполните 10 ударов с пестиком и процеживают через 100 мкм затем мкм клеточный фильтр 40 в центрифужную пробирку на 50 мл. Центрифуга в течение 10 мин медленнот 700 мкг при 4 ° C и удалить супернатант. Ресуспендируют ядерный осадок в 25 мл раствора в градиенте плотности сахарозы и слой Ядра клеток раствора на вершине 10 мл свежего раствора градиенте сахарозы в ультрацентрифуге трубки, полученного в разделе 3.1. Центрифуга при 13000 мкг в течение 1 ч при 4 ° С, используя качаться роторной трубы. После центрифугирования, отбросить супернатант и ресуспендируют ядер гранул в 1300 мкл ядер буфера для хранения в 1,5 мл трубки микроцентрифужных и этикетки PCM1. Удалить 300 мкл суспензии в новую пробирку микроцентрифужных и добавляют к 700 мкл свежего буфера для хранения ядер. Добавьте эту трубку в качестве контроля ISO. Добавить анти-PCM1 при конечной концентрации 8 мкг / мл в микроцентрифужных пробирку PCM1 и добавить кролика IgG-антитела контрольного изотипа в концентрации 8 мкг / мл в микроцентрифужных пробирку управления ISO. Инкубируют в течение ночи при температуре 4 ° С. После incubвания, центрифуге при 700 мкг в течение 10 мин, отбросить супернатант, заменить 1 мл свежего буфера для хранения ядра, ресуспендируют и центрифуга снова в течение 10 мин при 700 х г. Удалите супернатант и заменить 1 мл свежего буфера для хранения ядра. Добавить 1 мкл F (аb ') 2-фрагмент козьих антител против кроличьего IgG (H + L) антитела (FITC или эквивалентного зеленого флуоресцентного красителя), чтобы достичь конечной концентрации 2 мкг / мл. Примечание: Использование F (аb ') 2-фрагмент антитела, уменьшает вероятность возникновения неспецифического мечения клеток иммунной системы, включая макрофаги и В-клетки. Wrap трубы в алюминиевой фольгой для защиты от света и инкубировать в течение 1 часа при 4 ° С. 4. Обнаружение Edu инкорпорации Развести 10x концентрируют Edu реакционного буфера в соотношении 1:10 деионизированной водой. Центрифуга PCM-1 и управления меченых ядер суспензий ISO при 700 мкг в течение 10 мин, отбросить супернатант и ресуспендируют осадок ядер в 1 мл1% БСА / раствор PBS. Вымойте ядер гранул дважды. После последней промывки, отбросить супернатант и заменить 100 мкл Edu закрепителя. Обертка трубы в алюминиевой фольгой для защиты от света и инкубировать при комнатной температуре в течение 15 мин. Промыть образцы дважды 1 мл 1% БСА / раствор PBS, центрифугируют при 700 х г и инкубировать с 1x сапонина на основе раствора пермеабилизации при комнатной температуре в течение 15 мин. Приготовьте 1х EDU маркировки коктейль (таблица 2). Минимум 2 реакций необходимо будет не включать никакого контроля Edu. Добавить 500 мкл 1x EDU маркировки коктейль непосредственно к каждому образцу, уже содержащей ядра в 100 мкл 1х сапонина раствора на основе пермеабилизации и инкубировать при комнатной температуре в защищенном от света месте в течение 30 мин. Центрифуга при 700 мкг и ресуспендируют в 1 мл 1% BSA / PBS и повторить этот шаг еще два раза. Центрифуга при 700 мкг, отбросить супернатант и заменить 400 мкл раствора ДНК окрашивания (см Таблицу материалов). Название реагента 1. буфера для лизиса клеток 0,32 М сахарозы 10 мМ Трис-HCl (рН = 8) 5 мМ CaCl 2 5 мМ ацетат магния 2,0 мМ ЭДТА 0,5 мМ EGTA 1 мМ ДТТ 2.Sucrose Градиент Solution 2,1 М сахарозы 10 мМ Трис-HCl (рН = 8) 5 мМ ацетат магния 1 мМ ДТТ 3. Ядра буфер хранения (NSB) 0,44 М сахарозы 10 мМ Трис-HCl (рН = 7,2) 70 мМ KCl 10 мМ MgCl 2 1,5 мМ спермина Таблица 1: Ядра буферные изоляция ингредиенты Рецепт для всех буферов , используемых в разделе протокола 3 (ядер и диссоциации кардиомиоцитов ядер маркировки).. 5. Поток Cytometric Анализ Примечание: Выполните проточной цитометрии на изолированных ядрах , как описано ранее 7,11,15. Во- первых, создать сюжет , описывающий рассеяния вперед (FSC) и боковое рассеивание (SSC) , чтобы позволить дискриминацию ядер от мусора (Рисунок 2А). Создание сюжет дискриминировать одиночных ядер из агрегатов (Фигура 2В), откладывая 4 ', 6-диамидино-2-фенилиндол) (DAPI) область (3-405 / 450/50-А) по сравнению с высотой (33-405 / 450 / 50-Н). Обеспечить ТНАт-А 50 сигнал 3-405 / 450 / собрана на линейном масштабе, чтобы обеспечить правильное определение содержания ДНК. Создать график, описывающий 488/535/30-A (PCM-1) против SSC и отображать только события из синглетного ворот, созданного в 5.2, чтобы оценить PCM-1 выражение в популяции синглетного. Выполнить контрольный образец изотипа для того , чтобы установить ИКМ-1 положительные ворота (фиг.2с). Выполнить небольшое количество PCM1 меченый образец для проверки PCM1 выражения населения и литниковой позицию. Создать сюжет, описывающий SSC-A против 405/450/50-А (для обнаружения DAPI). На этом графике, отображение только синглетному PCM1, выражающих ядра, используя ворота, созданные в 5.3. Используйте этот участок , чтобы создать дополнительный иерархический ворота , который содержит только 2н ядра (рис 2D). Создать график, описывающий 488/535 / 30-A против 1-633 / 660/20-A, чтобы идентифицировать Edu маркировке PCM1, выражающей кардиомиоцитов населения. Показывать только ядра, которые являются синглет, P СМ1 + и 2н с использованием вышеуказанного стробирования. Запуск образца сердце от полного отсутствия контроля Эду установить Edu положительные ворота (рис 2E). Создать соответствующие ворота для Edu + клеток. Запись и количественного синглет, PCM-1, выражающий 2N ядер, которые включены EDU. Вычислить эту популяцию в процентах от общего синглета, PCM1 + ядер. Это обеспечит скорость нео-кардиомиоцитов ядер поколения, в течение 7-дневного периода ОБР импульса, как процент от общего количества ядер кардиомиоцитов. Примечание: В качестве DAPI связывания с молекулой ДНК является стехиометрическим интенсивность флуоресценции пропорциональна количеству ДНК. Определение плоидности на основе интенсивности / 450/50-А сигнала 405, как было описано выше 7. Для оценки плоидности в пределах общей популяции кардиомиоцитов использовать участок , установленный в 5.5 и стратегию , основанную на стробирования концентрации ДНК 7. /files/ftp_upload/53979/53979fig2.jpg "/> Рис . 2: стробирования стратегию количественной оценки кардиомиоцитов Edu включения и плоидности (A) Ядра подвергаются дискриминации от мусора на основе рассеяния вперед (FSC) и бокового рассеяния (SSC). (Б) Линейное ворота создается и синглетный популяция ядер определяется на основе на маркировке DAPI и 405/450/50 высоты против сигнала площади. (С) Флуоресцентные стробирования позволяет разделение ядер кардиомиоцитов (ПКМ-1-положительных) и некардиомиоцитов (PCM-1-отрицательных) ядер от сердечной ткани. (D) Интенсивность сигнала DAPI используется для определения концентрации ДНК и , таким образом , ядер плоидности кардиомиоцитов популяции PCM1 +. У мышей> 80% ядер кардиомиоцитов диплоидны (2n). (E) Люминесцентная стробирования позволяет разделить 2н, ядер кардиомиоцитов , которые включали Эду (PCM + / + EDU = 0,9%) от 2N, кардиомиоциты , которые не включены EDU (PCM + обученным), Все этапы подробно описаны в протоколе 5.0. Пример показан от MDX -. / У мышей на фоне C57 / BL10 Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.