Figures 2 – 4 illustrate the adaptation simulations for changes in the observer or the environment. Figure 2 compares the predicted appearance of Cezanne's Still Life with Apples for a younger and older observer who differ only in the density of the lens pigment28. The original image as seen through the younger eye (Figure 2a) appears much yellower and dimmer through the more densely pigmented lens (Figure 2b). (The corresponding shifts in the mean color and chromatic responses is illustrated in Figure 1c.) However, adaptation to the average spectral change discounts almost all of the color appearance change (Figure 2c). The original color response is almost completely recovered by the adaptation in the cones, so that subsequent contrast changes have negligible effect.
Figure 3 shows van Gogh's Irises filtered to simulate color appearance in a deuteranomalous observer, whose normal M photopigment is shifted in peak sensitivity to within 6 nm of the L photopigment28. Adaptation in the cones again adjusts for the mean stimulus chromaticity, but the L vs. M contrasts from the anomalous pigments are weak (Figure 3b), compressing the mechanism responses along this axis (Figure 1d). It has been suggested that van Gogh might have exaggerated the use of color to compensate for a color deficiency, since the colors he portrayed may appear more natural when filtered for a deficiency. However, contrast adaptation to the reduced contrasts predicts that the image should again "appear" very similar to the normal and anomalous trichromat (Figure 3c), even if the latter has much weaker intrinsic sensitivity to the L Vs. M dimension. Many anomalous trichromats in fact report reddish-greenish contrasts as more conspicuous than would be predicted by their photopigment sensitivities36,37.
Figure 4 shows the simulations for an environmental change, by simulating how the hazy image portrayed by Monet's Sunrise (Marine) might appear to an observer fully adapted to the haze (or to an artist fully adapted to his painting). Before adaptation the image appears murky and largely monochrome (Figure 4a), and correspondingly the mechanism responses to the image contrast are weak (Figure 1e). However, adaptation to both the mean chromatic bias and the reduced chromatic contrast (in this case to match the mechanism responses for typical outdoor scenes) normalizes and expands the perceived color gamut so that it is comparable to the range of color percepts experienced for well-lit outdoor scene (Figure 4b).
Finally, Figure 5 illustrates the two examples noted in section 6.2 of the protocol for using the model to study color vision. Figure 5a shows the Munsell Palette under adaptation to a lush or arid environment, while Figure 5b plots the shifts in the palette stimuli required to appear pure red, green, blue, or yellow, when the same observer is adapted to a range of different simulated environments. This range is comparable to measurements of the actual stimulus range of these focal colors as measured empirically in the World Color Survey29. Figure 5c instead shows how a set of embedded colors appear before or after adaptation to a Martian landscape. Adapting the set for the image led to significantly shorter reaction times for finding the unique colors in a visual search task29.
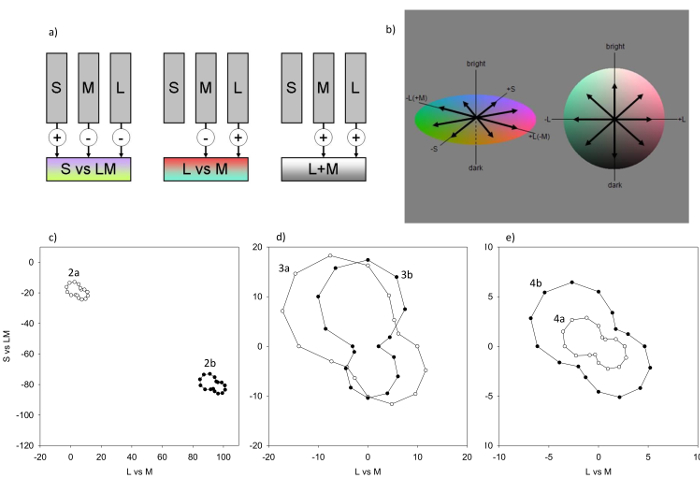
Figure 1: The Model. a) Responses are modeled for mechanisms with the sensitivities of the cones (which adapt to the stimulus mean) or postreceptoral combinations of the cones (which adapt to the stimulus variance. b) Each postreceptoral mechanism is tuned to a different direction in the color-luminance space, as indicated by the vectors. For the simulations 26 mechanisms are computed, which sample the space in 45° intervals (shown for the L Vs. M and S Vs. LM plane, and the L Vs. M and luminance plane). c) Responses of the mechanisms in the equiluminant (L Vs. M and S Vs. LM) plane to the images in the top and middle panel of Figure 2. Mean contrast responses are shown at 22.5° intervals to more fully portray the response distribution, though the model is based on channels at 45° intervals. In the original image (Figure 2a) the mean chromaticity is close to gray (0,0) and colors are biased along a bluish-yellowish axis. Increasing the lens density of the observer produces a large shift in the mean toward yellow (Figure 2b). d) Contrast responses for the images shown in Figure 3a & 3b. The cone contrasts in the original (Figure 3a) are compressed along the L Vs. M axis for the color deficient observer (Figure 3b). e) Contrast responses for the images shown in Figure 4a & 4b. The low contrast responses for the original image (Figure 4a) are expanded following adaptation, which matches the mean responses to the painting to the responses for a color distribution typical of outdoor natural scenes (Figure 4b). Please click here to view a larger version of this figure.

Figure 2: Simulating the Consequences of Lens Aging. Cezanne's Still Life with Apples (a) processed to simulate an aging lens (b) and adaptation to the lens (c). Digital image courtesy of the Getty's Open Content Program. Please click here to view a larger version of this figure.

Figure 3: Simulating Anomalous Trichromacy. van Gogh's Irises (a) simulating the reduced color contrasts in a color-deficient observer (b), and the predicted appearance in observers fully adapted to the reduced contrast (c). Digital image courtesy of the Getty's Open Content Program. Please click here to view a larger version of this figure.

Figure 4: Simulating Adaptation to a Low Contrast Environment. Monet's Sunrise (Marine). The original image (a) is processed to simulate the color appearance for an observer adapted to the low contrasts in the scene (b). This was done by adjusting the sensitivity of each mechanism's sensitivity so that the average response to the colors in the paintings is equal to the average response to colors measured for a collection of natural outdoor scenes. Digital image courtesy of the Getty's Open Content Program. Please click here to view a larger version of this figure.
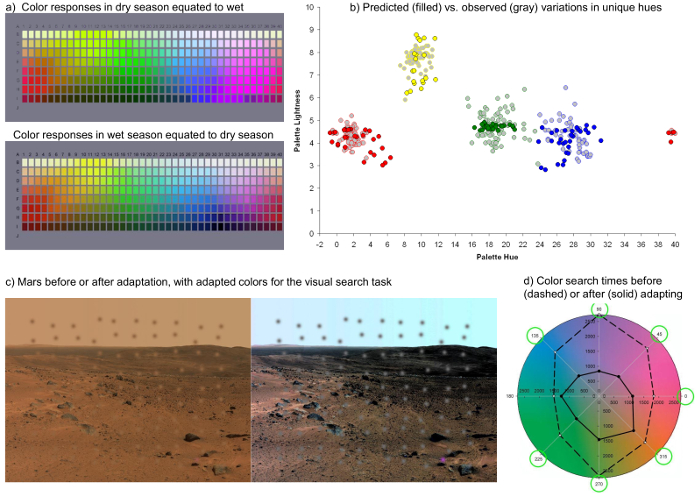
Figure 5. Using the Model to Examine Visual Performance. a) The Munsell palette rendered under adaptation to the colors of a lush or arid environment. b) Chips in the palette that should appear pure red, green, blue, or yellow after adaptation to a range of different color environments. Light-shaded symbols plot the range of average chip selections from the languages of the World Color Survey. c) Images of the surface of Mars as they might appear to an observer adapted to Earth or to Mars. Superimposed patches show examples of the stimuli added for the visual search task, and include a set of uniformly colored distractors and one differently-colored target. d) In the experiment search times were measured for locating the odd target, and were substantially shorter within the adapted Mars-adapted images. Please click here to view a larger version of this figure.
