Generation of Induced-pluripotent Stem Cells Using Fibroblast-like Synoviocytes Isolated from Joints of Rheumatoid Arthritis Patients
Summary
Here we describe a protocol for generating human induced-pluripotent stem cells from patient-derived fibroblast-like synoviocytes, using a lentiviral system without feeder cells.
Abstract
Mature somatic cells can be reversed into a pluripotent stem cell-like state using a defined set of reprogramming factors. Numerous studies have generated induced-Pluripotent Stem Cells (iPSCs) from various somatic cell types by transducing four Yamanaka transcription factors: Oct4, Sox2, Klf4 and c-Myc. The study of iPSCs remains at the cutting edge of biological and clinical research. In particular, patient-specific iPSCs can be used as a pioneering tool for the study of disease pathobiology, since iPSCs can be induced from the tissue of any individual. Rheumatoid arthritis (RA) is a chronic inflammatory disease, classified by the destruction of cartilage and bone structure in the joint. Synovial hyperplasia is one of the major reasons or symptoms that lead to these results in RA. Fibroblast-like Synoviocytes (FLSs) are the main component cells in the hyperplastic synovium. FLSs in the joint limitlessly proliferate, eventually invading the adjacent cartilage and bone. Currently, the hyperplastic synovium can be removed only by a surgical procedure. The removed synovium is used for RA research as a material that reflects the inflammatory condition of the joint. As a major player in the pathogenesis of RA, FLSs can be used as a material to generate and investigate the iPSCs of RA patients. In this study, we used the FLSs of a RA patient to generate iPSCs. Using a lentiviral system, we discovered that FLSs can generate RA patient-specific iPSC. The iPSCs generated from FLSs can be further used as a tool to study the pathophysiology of RA in the future.
Introduction
Pluripotent stem cells are the next-generation platform in various clinical and biological fields. They are a promising tool that can be used in disease modeling, drug screening, and regenerative medical therapy. Human Embryonic Stem Cells (hESCs) were mainly used to study and understand pluripotent cells. However, isolated by the destruction of the human blastocyst, hESCs are associated with several ethical concerns. In 2007, Dr. Shinya Yamanaka and his team reversed the cell programming process and developed stem cells from human adult somatic cells1,2. Therefore, unlike hESCs, induced-Pluripotent Stem Cells (iPSCs) can be generated from mature somatic cells, avoiding the ethical hurdles.
Usually, iPSCs are generated by the delivery of four exogenous genes: Oct4, Sox2, Klf4, and c-Myc. These Yamanaka factors are originally delivered using lentiviral and retroviral systems. The first iPSCs were derived from mouse somatic cells3. Afterwards, the technique was applied to human dermal fibroblasts1,2. Subsequent studies successfully generated iPSCs from various sources, such as urine4, blood5,6, keratinocytes7, and several other cell types. However, there are some somatic cells that have not been used in reprogramming, and screening of the reprogramming capabilities of various cell types from specific tissues in disease state, is still required.
Rheumatoid arthritis (RA) is a disease that can strike all joints and lead to autoimmune conditions in other organs. RA affects about 1% of adults in the developed world. It is a rather common disease and its incidence increases each year8. However, RA is hard to identify in the early stages and oncebone destruction occurs there is no treatment that can recover the damage. Moreover, drug efficacy differs from patient to patient, and it is hard to predict the medicine that is required. Therefore, the development of a drug-screening method is needed, and a cell material that can reflect the conditions of RA is required.
Fibroblast-like Synoviocytes (FLSs) are an active cellular participant in the pathogenesis of RA9,10. FLSs exist in the synovial intimal lining between the joint capsule and cavity, which is also referred to as the synovium. By supporting the joint structure and providing nutrients to the surrounding cartilage, FLSs usually play a crucial role in joint function and maintenance. However, FLSs in RA have an invasive phenotype. RA FLSs have a cancer-like phenotype, eventually destroying the surrounding bone by infinite proliferation10. With this unique characteristic, FLSs can be used as a promising material that can reflect the pathobiology of RA. Yet, these cells are rarely produced, and the cell phenotypes alter as the cells go through several passages in in vitro conditions. Therefore, it can be complicated to use RA FLSs as a tool that can represent the patient's condition.
Theoretically, RA patient-derived iPSCs (RA-iPSCs) can become an ideal tool for drug screening and further research. Generated iPSCs have self-renewal ability and can be maintained and expanded in vitro. With pluripotency, these cells can be differentiated into mature chondrocyte and osteocyte lineages, which can contribute cell material for specific research in RA and other bone-related diseases11.
In this study, we demonstrate how to isolate and expand FLSs from a surgically removed synovium, and how to generate RA-iPSCs from FLSs using lentiviruses containing Yamanaka factors.
Protocol
Ethics Statement: This study protocol was approved by the institutional review board of The Catholic University of Korea (KC12TISI0861).
1. Synoviocyte Isolation and Expansion
- Synoviocyte Isolation
- Sterilize two pairs of surgical scissors and one pair of forceps.
- Transfer the synovial tissue to a 100 mm dish and wash with 5 ml of phosphate-buffered saline (PBS) containing 1% penicillin/streptomycin.
- Cut off the yellowish fat tissue and bone residues. Transfer the trimmed tissue to a well of a 6-well plate and add 5 ml of Dulbecco's modified Eagle's medium (DMEM) with 20% Fetal bovine serum (FBS).
- Chop the tissues with the scissors until the pieces are small enough to penetrate a disposable pipette.
- Transfer the tissue-containing media to a 50 ml conical tube. Harvest the remaining material by adding 5 ml of DMEM with 20% FBS to the 6-well plate and then transfer to the tube.
- Thaw collagenase on ice. Add collagenase to a final concentration of 0.01% and seal the tube with parafilm. Incubate in a water bath at 37 °C with shaking for 4 hr.
- After incubation, fill the tube with DMEM with 20% FBS, until the total volume is 50 ml and centrifuge at 300 x g, RT for 10 min.
- Remove the supernatant without disturbing the pellet and add 40 ml of media to resuspend the pellet.
- Repeat steps 1.1.10-1.1.11.
- Resuspend the pellet in 25 ml of DMEM with 20% FBS and wait for the large clumps of tissue to sink to the bottom.
- Transfer the supernatant to a 100 mm dish and incubate at 37 °C in 5% CO2 for 14 day.
- Synoviocyte Maintenance and Expansion
- Discard the used media from the plate and wash the cells with 5 ml of PBS.
- Add 1 ml PBS/1 mM EDTA and incubate at 37 °C in 5% CO2 for 2 min.
- Tap the dish gently and transfer the cells to a 15 ml conical tube. Centrifuge the cells at 250 x g, RT for 2 min.
- Remove the supernatant without disturbing the pellet and resuspend the pellet in 30 ml of DMEM with 20% FBS.
- Transfer the cells to 3 x 100 mm dishes, without leaving any visible leftover material.
- Replace the media with fresh media every 3 d. Split the cells at 80% confluency using 1 ml PBS/1 mM EDTA. Maintain until passage 3 before use. Divide each dish of cells into 3 dishes in every split.
NOTE: After reaching passage 3, cells that are not going to be used immediately can be frozen.
2. Reprogramming FLSs Using Lentiviruses-encoding Yamanaka Factors
- Transduction (D0)
- Seed 3 × 104 cells per well of a 6-well plate in growth media (500 ml of DMEM supplemented with 10% FBS and 1% penicillin/streptomycin). Incubate the cells O/N at 37 °C in 5% CO2.
- The following day, remove one vial of lentivirus containing 4 Yamanaka factors: Oct4, Klf4, Sox2 and c-Myc from the freezer and thaw at 4 °C. Note: Lentivirus was produced by the procedure described in our previous study11.
- While thawing the virus, change the media to FLS growth media (20% FBS plus antibiotics in DMEM) containing 10 µg/ml hexadimethrine bromide and 50 µg/ml ascorbic acid.
- After changing the media, add 30 µl of lentivirus to the cells and mix gently. To improve infection, centrifuge the plate at 680 x g, 35 °C for 30 min.
- After centrifugation, incubate the cells at 37 °C in 5% CO2.
- Maintenance Until Reprogramming is Visible
- For 3 day, replace the media daily with FLS growth media containing 0.1 mM sodium butyrate and 50 µg/ml ascorbic acid.
- The next day, replace the media with a mixture of FLS growth media and iPSC media (1:1 ratio) containing 0.1 mM sodium butyrate and 50 µg/ml ascorbic acid.
Note: The components of the iPSC media is given in the materials/equipment list.
- Splitting Cells for Colony Formation
- Prepare a vitronectin-coated 6-well plate.
- Add 60 µl vitronectin to 6 ml PBS without Ca2+ and Mg2+. Put 2 ml of mixture into each wells and incubate in RT for at least 1 hr. Note: The working concentration of vitronectin is 5 µg/mL.
- On D5, wash the cells with PBS.
- Add 1 ml PBS/1 mM EDTA to detach the cells and incubate at 37 °C, 5% CO2 for 2 min.
- Harvest the cells and centrifuge at 250 x g, RT for 2 min.
- Split the cells at 3 different ratios (1:3, 1:6, and 1:9) to achieve different confluencies. Add 900 µl of media to the cell pellet and resuspend. Add 300, 150, and 100 µl of the cell mixture per well of a 6-well plate to achieve a ratio of 1:3, 1:6, and 1:9, respectively.
- Replace the media daily with iPSC media until colonies appear. Colonies will appear after about D18. Note: At this stage, iPSC colonies co-exist with the non-reprogrammed FLSs.
- Prepare a vitronectin-coated 6-well plate.
- Colony picking
- Prepare a 48-well vitronectin-coated plate by adding 500 µl vitronectin to the wells, and incubate at RT for at least 1 hr.
- Place the microscope on a clean bench, and remove the 6-well plate from the incubator.
- Remove the vitronectin solution from the 48-well plate and add 500 µl iPSC media supplemented with 10mM Rho-associated, coiled-coil containing protein kinase (ROCK) inhibitor.
- Using a 10p pipette tip, cut around the colony. Transfer the picked colony to one well of the 48-well plate.
- After picking several colonies, incubate the cells at 37 °C, 5% CO2.
- Maintain the cells until the colonies are big enough for transfer. Note: We usually spilt the cells when the colony gets out of the visible field of the microscope, when viewed at 100X magnification.
3. Immunofluorescence Staining
- Cell Preparation
- Place a sterile 18 mm cover glass into a 12-well plate.
- Add 1 ml of PBS to cool and rinse the cover glass.
- Replace with 1 ml of a 10 µg/ml vitronectin solution.
- Incubate the plate at RT for at least 1 hr.
- Discard the vitronectin solution and plate iPSCs in the vitronectin-coated 12-well plate and culture for 7 days at 37 °C, 5% CO2, changing the media daily.
- Cell Staining
- Discard the culture media and wash the cells with PBS once.
- Fix the cells in 0.4% paraformaldehyde (PFA) for 30 min at RT.
- Permeabilize the cells with 0.1% Triton X-100 for 5 min at RT.
- Remove the permeabilization solution and block with PBS containing 2% bovine serum albumin (BSA) for 30 min at RT.
- Dilute the antibodies in PBS containing 2% BSA according to Table 1. Incubate the cells with the primary antibodies for 2 hr at RT.
- Add the secondary antibody (diluted 1:200) and incubate the cells for 1 hr at RT, avoiding light.
- Treat the cells with 1 µl/mL DAPI for 10 min.
- Place the cover glass on top of the slide glass with antifade reagent and incubate at RT for 24 hr, avoiding light.
- Verify expression with a fluorescence microscope.
4. Real-time Polymerase Chain Reaction (RT-PCR)
- Extract mRNA from the cell pellet using the guanidinium thiocyanate-phenol-chloroform extraction method11.
- Amplify cDNA from 2 µg of total mRNA using reverse transcription11.
- Mix the components required for PCR using 2 µl of cDNA template11.
- Perform RT-PCR and verify the results by gel electrophoresis11.
5. Alkaline Phosphatase (AP) Staining
- Culture iPSCs for 5-7 days at 37 °C, 5% CO2 prior to staining.
- Aspirate the media and fix the cells with 4% PFA for 1 min.
- Discard the fixative and rinse the cells with 1X rinse buffer.
- Prepare the reagents for AP staining. Mix the reagents in the following ratio: Fast Red Violet : Naphthol AS-BI phosphate solution : water = 2:1:1.
- Incubate the cells with the staining solution at RT for 15 min, avoiding light.
- Discard the staining solution and rinse the cells with rinse buffer.
- Cover the cells with PBS to prevent drying and verify expression using a bright-field microscope.
Representative Results
In this study, we describe a protocol to generate iPSCs from FLSs using a lentiviral system. Figure 1A shows a simple scheme of the FLS isolation protocol. Following surgical removal of the synovium, the tissue was chopped into small pieces using surgical scissors. Collagenase was added to isolate the cells from the clumps of tissue. Cells were incubated for 14 days before further processing. Figure1B shows the morphology of the isolated FLSs. Cells were maintained for 3 passages before use. FLSs share a similar characteristic with general fibroblasts. Our isolated FLSs expressed the fibrotic markers, Vimentin and Fibronectin. The isolated RA FLSs also showed low expression of the macrophage-like synoviocyte marker, CD68 (Figure 1C).
FLSs were harvested and transduced with lentiviruses containing 4 Yamanaka factors for reprogramming. After splitting the cells at various ratios (D7), small colonies started to appear. Visible colonies, as shown in Figure 2A, appeared at D8-11. Colonies can be picked from this point. Figure 2B shows an image of a picked and amplified colony.
iPSCs were expanded until passage 5-10 and then used in various assays. Undifferentiated ESCs are characterized by a high level of AP. Cells were stained for AP to confirm the undifferentiated state. RA-iPSCs expressed AP (Figure 2C), indicating they are undifferentiated. Pluripotent marker expression was examined (Figure2D,E). The expression of pluripotent markers such as Oct3/4, Sox2, Nanog, Lin28, DPPB5 and TDGF1 was confirmed using RT-PCR (Figure 2D). The expression of Oct3/4, Sox2 was also confirmed by immunofluorescence analysis with additional markers such as SSEA4, TRA-1-60, TRA-1-81 and Klf4. TRA-1-60, which is currently thought to be the most important iPSC marker, was highly expressed in our generated iPSCs.
For further analysis, we performed karyotyping and teratoma assay. RA-iPSCs showed a normal chromosomal pattern of 44 + XY (Figure 3A). 12 weeks-post injection of RA-iPSCs into SCID mice, teratomas had formed and displayed diverse tissues, such as gland, adipose tissue and blood vessels (Figure 3B). The germ layer differentiation was also confirmed through immunofluorescence staining. Ectoderm lineage cells were positively stained for Otx2. Mesodermal cells expressed brachyury, and endoderm was confirmed by positive staining of SOX17.
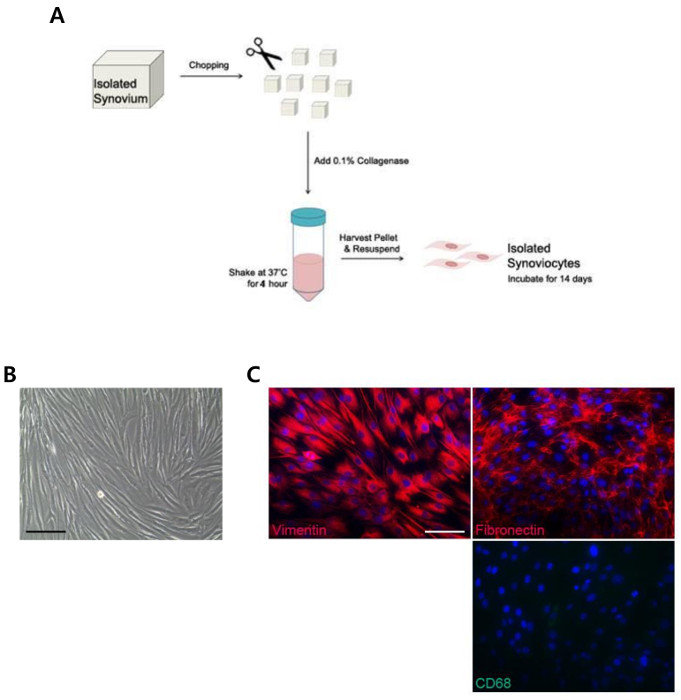
Figure 1: Protocol for FLS Isolation from a RA Patient. (A) A simple diagram of the method used for synoviocyte isolation. (B) Morphology of isolated FLSs. (C) Fluorescence microscopy image of FLSs stained with fibrotic markers vimentin, fibronectin and a macrophage-like synoviocyte marker, CD68. All Scale bars = 200 µm. Please click here to view a larger version of this figure.
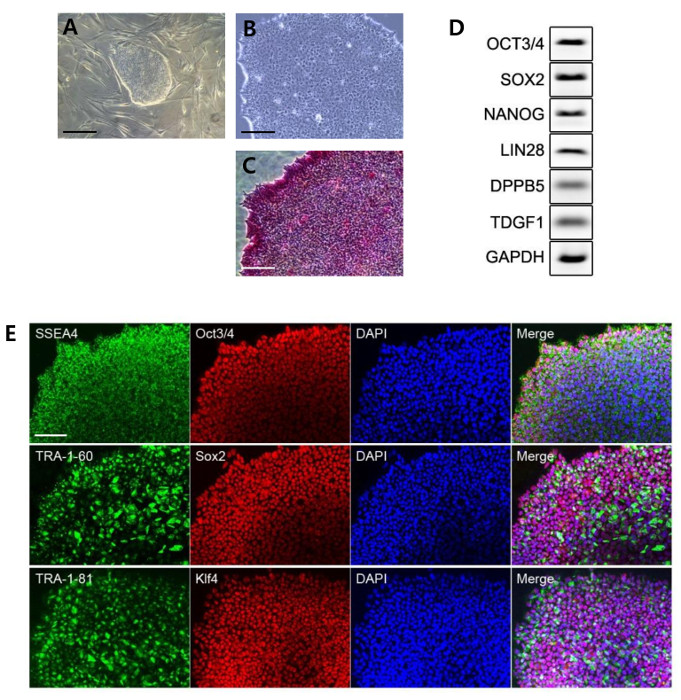
Figure 2: Generation of iPSCs from FLSs Isolated from a RA Patient. (A) Bright-field image of an iPSC colony before picking. (B) Image of a colony after picking. (C) Colony stained for AP. (D) PCR analysis of pluripotent markers. (E) Fluorescence microscopy image of iPSCs. The generated iPSCs expressed all the pluripotent markers. All Scale bars = 200 µm. Please click here to view a larger version of this figure.
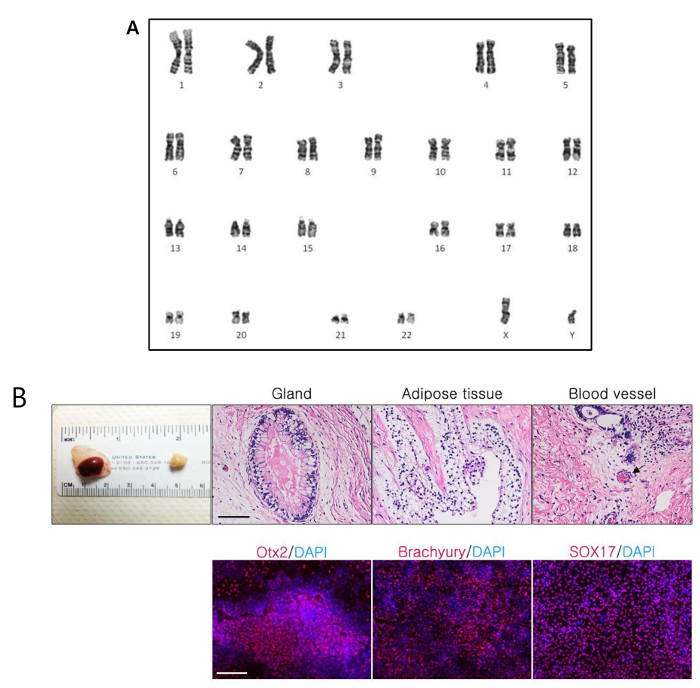
Figure 3: Karyotype Analysis and Teratoma Assay. (A) High resolution G-banded images showing normal karyograms of iPSCs. The image represents a 46XY normal diploid male chromosomal content. (B) Results of teratoma assays and immunofluorescence staining. All Scale bars = 200 µm. Please click here to view a larger version of this figure.
Discussion
Before the discovery of iPSCs, scientists mainly used ESCs to study stem cell biology and other cell lineages through differentiation. However, ESCs originate from the inner mass of a blastocyst, which is an early-stage embryo. To isolate ESCs, destruction of the blastocyst is inevitable, raising ethical issues that are impossible to overcome. Moreover, although ESCs have stemness characteristics and pluripotency, they cannot be obtained from individuals and are sometimes not an ideal tool for personalized analysis and disease screening.
In 2007, Takahashi et al. generated iPSCs from human fibroblasts2. Theoretically, unlike ESCs, iPSCs can be generated from any adult somatic cells. With this advantage, iPSCs are thought to be the ideal tool for auto-cell transplantation. Also, with the concept of epigenetic memory, iPSCs are thought to be the ideal cell material for the simulation of pathogenic conditions, i.e. disease modeling. There have been many studies done using various types of cells such as blood cells, urine cells and more. Yet, there are many cell types, such as FLSs, in which reprogramming has not been conducted.
Arthritic diseases are major immune disorders that can cause permanent disability. RA is caused by chronic inflammation in the joints, eventually resultimg in bone and cartilage damage. It is hard to cure or reverse the damage especially because cartilage cannot regenerate in vivo. Therefore regenerative medicine using iPSCs are a new critical tool for the cure of RA. The bone and cartilage loss is also resulted in pannus formation. Pannus is a horn-like structure that can be seen in the histological image of the joint. The pannus is made by FLSs that proliferate limitlessly like cancer cells. Therefore, it is thought that the FLSs can reflect the pathological characteristics of the disease. In this study, we used patient FLSs to generate iPSCs.
FLSs are the major cell type that contribute to the pathogenesis of RA. As a cell that is mostly exposed to the inflammatory environment inside the synovial joint, our group thought that it can be used as a material that reflects the patient's disease condition. Therefore, using the earliest reprogramming method, we attempted to generate RA-specific iPSCs from FLSs, using the delivery of the 4 Yamanaka factors – Oct3/4, Sox2, Klf4 and c-Myc – by lentivirus. FLSs were isolated from the removed synovium. The trimming of synovium was critical when isolating FLSs. Bone and fat residues can make it more complicated to obtain pure FLSs. Also, the use of collagenase is inevitable for FLS isolation. It is important to use cells between passages 3-8. When FLSs reach passage 3, the Yamanaka factor containing lentiviruses are generated following the procedure mentioned in our earlier work11. Using the produced lentiviruses, we successfully generated RA FLS-derived iPSCs (RA-iPSCs). Pure iPSC clones were generated by the colony picking method. The RA-iPSCs expressed all the pluripotent transcription markers and had a normal karyotype. Furthermore, the RA-iPSCs were able to differentiate into all three germ layers according to the teratoma assay. Also, we have confirmed that the RA-iPSCs showed more mineralization when differentiated into osteogenic lineages in vitro (data not shown) 11.
However, there are some limitations to this method. Lentiviruses require genomic integration for reprogramming. Klf-4 and c-Myc are oncogenes and these two Yamanaka factors can facilitate tumor growth in vivo. Therefore, it may not be ideal to use these factors to generate materials for clinical applications. Furthermore, FLSs and skin fibroblasts are not difficult to obtain in clinics. As mentioned in various reports, skin fibroblasts (i.e. dermal fibroblasts) can only be obtained by punch biopsies. FLSs can be isolated only by an invasive surgical procedure and this surgery is performed on patients with severe hyperplasia who have undergone knee surgery. Therefore, there is no need to use FLSs when reprogramming a non-RA iPSC. Additionally, the process by which fibroblasts are prepared for reprogramming is difficult and time-consuming. FLSs share the same shortcomings as skin fibroblasts. Therefore, cell sources that are easier to handle and obtain are required.
For this reason, researchers have begun to search for an alternative cell source. A currently used material is blood cells. It is easy to draw blood and the isolation process is relatively simple and fast. Furthermore, there is a shift from the use of lentiviruses to tools that do not require genome-integration, such as Sendai viral systems, small molecules, and episomal plasmids.
Although FLSs cannot be used as a material for diverse individuals, it is still a great material when generating RA-iPSCs for research purposes. In the future, we are looking to generate a RA patient FLS-derived iPSC bank. RA patients individually show diverse reactions to drugs that are used for treatment. With several lines of RA iPSCs, we are hoping to generate a drug-screening cell-bank for RA patients. By screening all the drugs on each cell line, we may be able to predict which drug will work on which individual patient.
In conclusion, this protocol describes the application of iPSC technology to rheumatology. With this protocol, iPSCs can be generated from RA patient-derived FLSs, and the generated iPSCs have the required characteristics. These iPSCs can be used in clinical research, drug screening, disease modeling, and regenerative medicine for further investigations of the biology of RA.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was supported by the Research Program funded by the Korea Centers for Disease Control and Prevention (HI13D2188).
Materials
| 100mm Dish | TPP | 93100 | |
| 6-well Plate | TPP | 92006 | |
| 50 mL Cornical Tube | SPL | 50050 | |
| 15 mL Cornical Tube | SPL | 50015 | |
| 10 mL Disposable Pipette | Falcon | 7551 | |
| 5 mL Disposable Pipette | Falcon | 7543 | |
| 12-well Plate | TPP | 92012 | |
| FLS Isolation Materials | |||
| Surgical Scissors | |||
| Surgical Forcep | |||
| DPBS | Life Technologies | 14190-144 | |
| DMEM | Life Technologies | 11995-073 | |
| Penicilin Streptomycin | Sigma Aldrich | P4333 | |
| Fetal Bovine Serum | Life Technologies | 16000-044 | |
| Collagenase | Sigma Aldrich | C6885-100MG | |
| Parafilm | Sigma Aldrich | 54956 | |
| PBS/1 mM EDTA | Life Technologies | 12604-039 | |
| iPSC Generation Materials | |||
| DMEM | Life Technologies | 11885 | |
| MEM Non-Essential Amino Acids Solution (100X) | Life Technologies | 11140-050 | |
| β-Mercaptoethanol | Sigma Aldrich | M3148 | |
| Polybrene | Chemicon | TR-1003-G | |
| Penicilin Streptomycin | Life Technologies | P4333 | |
| Fetal Bovine Serum | Life Technologies | 16000-044 | |
| DPBS | Life Technologies | 14190-144 | |
| Lentivirus | |||
| DMEM/F12, HEPES | Life Technologies | 11330-057 | iPSC media ingredient (500 mL) |
| Sodium Bicarbonate | Life Technologies | 25080-094 | iPSC media ingredient (Conc.: 543 μg/mL) |
| Sodium Selenite | Sigma Aldrich | S5261 | iPSC media ingredient (Conc.: 14 ng/mL) |
| Human Transfferin | Sigma Aldrich | T3705 | iPSC media ingredient (Conc.: 10.7 μg/mL) |
| Basic FGF2 | Peprotech | 100-18B | iPSC media ingredient (Conc.: 100 ng/mL) |
| Human Insulin | Life Technologies | 12585-014 | iPSC media ingredient (Conc.: 20 μg/mL) |
| Human TGFβ1 | Peprotech | 100-21 | iPSC media ingredient (Conc.: 2 ng/mL) |
| Ascorbic Acid | Sigma Aldrich | A8960 | iPSC media ingredient (Conc.: 64 μg/mL) |
| Polybrene | Chemicon | TR-1003 | |
| Sodium Butyrate | Sigma Aldrich | B5887 | |
| Vitronectin | Life Technologies | A14700 | |
| ROCK Inhibitor | Sigma Aldrich | Y0503 | |
| Guality Control Materials | |||
| 18 mm Cover Glass | Superior | HSU-0111580 | |
| 4% Paraformaldyhyde | Tech & Innovation | BPP-9004 | |
| Triton X-100 | BIOSESANG | 9002-93-1 | |
| Bovine Serum Albumin | Vector Lab | SP-5050 | |
| Anti-SSEA4 Antibody | Millipore | MAB4304 | |
| Anti-Oct4 Antibody | Santa Cruz | SC9081 | |
| Anti-TRA-1-60 Antibody | Millipore | MAB4360 | |
| Anti-Sox2 Antibody | Biolegend | 630801 | |
| Anti-TRA-1-81 Antibody | Millipore | MAB4381 | |
| Anti-Klf4 Antibody | Abcam | ab151733 | |
| Alexa Fluor 488 goat anti-mouse IgG (H+L) antibody | Molecular Probe | A11029 | |
| Alexa Fluor 594 goat anti-rabbit IgG (H+L) antibody | Molecular Probe | A11037 | |
| DAPI | Molecular Probe | D1306 | |
| Prolong gold antifade reagent | Invitrogen | P36934 | |
| Slide Glass, Coated | Hyun Il Lab-Mate | HMA-S9914 | |
| Trizol | Invitrogen | 15596-018 | |
| Chloroform | Sigma Aldrich | 366919 | |
| Isoprypylalcohol | Millipore | 109634 | |
| Ethanol | Duksan | 64-17-5 | |
| RevertAid First Strand cDNA Synthesis kit | Thermo Scientfic | K1622 | |
| i-Taq DNA Polymerase | iNtRON BIOTECH | 25021 | |
| UltraPure 10X TBE Buffer | Life Technologies | 15581-044 | |
| loading star | Dyne Bio | A750 | |
| Agarose | Sigma-Aldrich | 9012-36-6 | |
| 1kb (+) DNA ladder marker | Enzynomics | DM003 | |
| Alkaline Phosphatase | Millipore | SCR004 |
References
- Yu, J., et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 318 (5858), 1917-1920 (2007).
- Takahashi, K., et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131 (5), 861-872 (2007).
- Takahashi, K., Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 126 (4), 663-676 (2006).
- Zhou, T., et al. Generation of human induced pluripotent stem cells from urine samples. Nat Protoc. 7 (12), 2080-2089 (2012).
- Haase, A., et al. Generation of induced pluripotent stem cells from human cord blood. Cell Stem Cell. 5 (4), 434-441 (2009).
- Loh, Y. H., et al. Generation of induced pluripotent stem cells from human blood. Blood. 113 (22), 5476-5479 (2009).
- Aasen, T., et al. Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes. Nat Biotechnol. 26 (11), 1276-1284 (2008).
- Scott, D. L., Wolfe, F., Huizinga, T. W. Rheumatoid arthritis. Lancet. 376 (9746), 1094-1108 (2010).
- Chang, S. K., Gu, Z., Brenner, M. B. Fibroblast-like synoviocytes in inflammatory arthritis pathology: the emerging role of cadherin-11. Immunol Rev. 233 (1), 256-266 (2010).
- Bartok, B., Firestein, G. S. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunol Rev. 233 (1), 233-255 (2010).
- Lee, J., et al. Generation of disease-specific induced pluripotent stem cells from patients with rheumatoid arthritis and osteoarthritis. Arthritis Res Ther. 16 (1), R41 (2014).
