All animal experiments described below were approved by the Committee for Medical Ethics and the use of Experimental Animals at the University of Antwerp (file number 2012-42).
1. Tissue Preparation of Jejunal and Colonic Afferent Nerves
- Preparation of the jejunal afferent nerve
- Perform rodent euthanasia of the adolescent or adult rodent that has been approved prior to the experiment by the local Ethical Committee (e.g., terminal sedation followed by cardiac puncture, cervical dislocation, etc.).
NOTE: We used cervical dislocation to sacrifice the animals thus resulting in experiments without the need for further anesthesia or post-surgical care as tissues are further processed in vitro.
NOTE: Age has been shown to attenuate mesenteric afferent mechanosensory functions 30, we therefore advise researchers to adhere to a specific age-group for the duration of a single experiment. - Place the sacrificed laboratory animal in the supine position and perform an abdominal midline incision through the skin and abdominal muscle layer using a scalpel, extending from the xyphoid process until the pubic bone.
- Bathe the abdominal cavity with cold Krebs solution in order to prevent the intra-abdominal tissues from drying out (Krebs composition: 120.03 mM NaCl, 6.22 mM KCl, 1.57 mM NaH2PO4, 15.43 mM NaHCO3, 1.21 mM MgSO4, 11.52 mM D-glucose and 1.52 mM CaCl2).
- Rapidly excise the entire jejunum using sharp scissors by excising approximately 20 cm of the small bowel starting immediately distally of the duodenojejunal flexure, taking care not to damage surrounding structures and keeping the bowel's mesentery, which contains jejunal blood vessels and afferent nerves, intact.
NOTE: For the mere jejunal dissection in the abdominal cavity, one does not need to use a stereomicroscope, as this can be easily visualized with the naked eye. - Place the excised jejunum in ice-cold Krebs solution and keep on ice, while oxygenating the Krebs solution continuously with carbogen (95% O2, 5% CO2).
- Cut the jejunum with sharp scissors in approximately 3-cm long loops. Observe the mesenteric bundle containing the vessels and splanchnic nerve somewhere near the center of the respective loop.
- Flush each segment with Krebs solution using a blunt catheter to remove luminal contents and chyme as these contain digestive enzymes that will accelerate the deterioration of the tissue sample in vitro.
NOTE: Take care not to damage the lumen of the loop during flushing, as the destruction of the villi will result in the release of mediators that can alter the afferent nerve activity. - Identify the segment to measure the afferent nerve activity (e.g., the first jejunal segment distally of the ligament of Treitz or the duodenojejunal flexure), and place this in a purpose-built recording chamber coated with a silicone elastomer layer.
NOTE: The beginning of the jejunum is anatomically defined as the part of the small bowel where the ligament of Treitz crosses the small bowel, also called the duodenojejunal flexure.
NOTE: Cover the bottom of the recording chamber with a thin silicone elastomer layer well in advance before the start of the experiment. The preparation of this elastomer layer should be performed according to the manufacturer's instructions 1. - Perfuse the chamber constantly with warm, carbogenated Krebs solution at a rate of 10 ml/min and keep the Krebs' temperature in the recording chamber constant at 34 °C.
- Mount the jejunal segment in the organ bath so that the oral end is connected to the syringe driver providing luminal flow and the aboral end connects to the outflow. Slightly stretch the segment but take care not to exert excessive tension. Attach both ends firmly using 4/0 silk ligatures to the in- and outflow ports.
- Attach the syringe driver to the oral end, and perfuse the jejunal segment intraluminally with Krebs solution (non-oxygenated, ambient temperature) at a rate of 10 ml/hr.
- Pin the mesentery of the mounted intestinal segment flat against the silicone elastomer bottom layer, using insect pins. Stretch the mesentery out in order to optimize visualization of the mesenteric bundle; do not exert strain on the bundle or the jejunum.
- Perform a test ramp distension (vide infra) by closing the output port until the intraluminal pressure of the intestinal segment reaches 60 mmHg, in order to verify that no intraluminal Krebs solution is leaking from the mounted segment. Observe a smooth rise in intraluminal pressure without interruptions.
- Observe small contractions of the segment (peristaltic waves) during the initial distension phase. If required, block peristaltic activity by adding 1 µM of the L-type calcium channel blocker nifedipine to the Krebs solution.
NOTE: Adding 1 µM of atropine to the Krebs solution in addition to nifedipine, will completely paralyze the intestinal segment. We however have no personal experience with the use and effect of atropine on afferent nerve recording. - Under a stereomicroscope, gently start to peel off the fat tissue from the mesentery by gently tugging it with two small tweezers, taking care not to damage the vessels and afferent nerve that lie buried in the fat tissue.
- Start at a remote distance from the jejunum, and expose both blood vessels in the mesenteric bundle.
- Observe the afferent jejunal nerve in between both vessels as a thin, white thread encapsulated in adipose tissue. Only dissect more proximally towards the jejunum by gently peeling the fat tissue away using tweezers when the initial identification of both mesenteric vessels and/or the afferent nerve is difficult.
- Dissect the jejunal mesenteric nerve of the segment free over a distance of several millimeters, by removing the adipose tissue adherent to the nerve.
- Transect the nerve using sharp tissue scissors. If required, peel off the remaining fat and connective tissue, as well as the epineuronal sheath by gently tugging it with the small tweezers.
- Using a micromanipulator, lower the tip of the suction electrode, connected to a syringe with plunger, into the organ bath; then, by manipulating the plunger, gently aspirate some Krebs solution from the organ bath so that the tip of the electrode is submerged in the Krebs solution (Figure 1). Ensure that the Krebs solution covers the wire electrode inside the suction electrode.
NOTE: Prepare the borosilicate glass suction capillary that contains the wire electrode prior to the start of the experiment using a pipette puller. - Position the tip of the suction electrode immediately next to the transected afferent nerve strand and draw the transected nerve strand into the capillary over its entire length.
- Maneuver the tip of the electrode towards some adipose tissue and aspirate this into the glass capillary while firmly aspirating with the plunger, thereby mechanically 'sealing' the nerve in the capillary from the contents of the organ bath.
- Verify the recording of afferent nerve activity using the data acquisition system, e.g., by performing a ramp distension-induced increase in afferent firing (vide infra). Following the isolation of the nerve into the suction electrode, stabilize the preparation for 15 min in order to obtain a steady state spontaneous afferent nerve activity before performing the actual experiments.
- To perform ramp distension, distend the intestinal segment by closing the output port, leading to gradual rise in pressure in the intestinal segment (up to 60 mmHg). Only perform the desired experimental protocol when three consecutive ramp distensions (with a 15 min interval) yield a reproducible multi-unit discharge (Figure 2).
- Perform rodent euthanasia of the adolescent or adult rodent that has been approved prior to the experiment by the local Ethical Committee (e.g., terminal sedation followed by cardiac puncture, cervical dislocation, etc.).
- Preparation of the lumbar splanchnic afferent (colonic afferent nerve)
NOTE: The dissection of colonic afferent nerves requires a more detailed dissection. Deviations from the former 'jejunal' protocol are listed below:- Euthanize the animal by means of a humane method, place it in the supine position, perform a midline laparotomy and excessively pour ice-cold Krebs solution in the abdominal cavity. NOTE: Krebs composition: 118 mM NaCl, 4.75 mM KCl, 1 mM NaH2PO4, 22 NaHCO3, 1.2 mM MgSO4, 11 mM D-glucose and 2.5 mM CaCl2, 3 µM indomethacin.
NOTE: Note the altered composition of the Krebs solution; indomethacin is added to the solution in order to prevent alterations of afferent nerve activity by prostaglandins. - Discard the adipose tissue, bladder and internal genitalia, and gently shift the small bowel to one side in the abdominal cavity. Perform an extended prelevation of the distal part of the colon with intact mesenteric nerves from the abdomen.
NOTE: Important landmarks that may be included in this dissection to aid further preparation of the tissue include the abdominal aorta and vena cava, the left kidney and the pelvic floor musculature.
NOTE: During the dissection, take care not to exert traction on the connecting tissue between colon and the abdominal aorta, as this region contains the lumbar splanchnic nerves. - Transfer the tissue segment into the silicone elastomer lined recording chamber. Use the left renal artery, which originates from the abdominal aorta, as a starting point. Follow the abdominal aorta in the aboral direction, encounter the superior mesenteric artery together with the celiac and superior mesenteric ganglion. Finally, arrive at the connecting tissue between the distal part of the colon and the aorta, wherein the nerve of interest is located.
- Identify the inferior mesenteric artery originating from the abdominal aorta. The lumbar splanchnic afferent nerve can be identified at the base of the inferior mesenteric artery; it runs parallel to the artery (Figure 3).
- Following the transection of the nerve, gently peel off the surrounding connective tissue sheath and tease the nerve into several fine threads. Make sure to keep a safe distance from the colon.
- Draw one of these single threads into the suction electrode, 'seal' the capillary with surrounding adipose tissue as previously described and perform the desired experimental protocol.
- Discard any superfluous tissue that is present in the organ bath (e.g., kidneys, abdominal vessels, muscle tissue), as these may disturb the afferent signal.
NOTE: Nifedipine (1µM) can be added to the Krebs solution in case the afferent nerve signal is disturbed by spontaneous intestinal movement due to smooth muscle cell contractions.
NOTE: Drugs can be administered at three different sites: 1) serosally by dissolving the desired compound into the Krebs that perfuses the recording chamber, 2) directly into the organ bath while temporarily stopping the perfusion or 3) intraluminally by dissolving the compound of interest into the Krebs solution in the syringe drive. Desensitization of the afferent nerve can occur when cumulative dosages of a compound are administered too fast consecutively.
- Euthanize the animal by means of a humane method, place it in the supine position, perform a midline laparotomy and excessively pour ice-cold Krebs solution in the abdominal cavity. NOTE: Krebs composition: 118 mM NaCl, 4.75 mM KCl, 1 mM NaH2PO4, 22 NaHCO3, 1.2 mM MgSO4, 11 mM D-glucose and 2.5 mM CaCl2, 3 µM indomethacin.
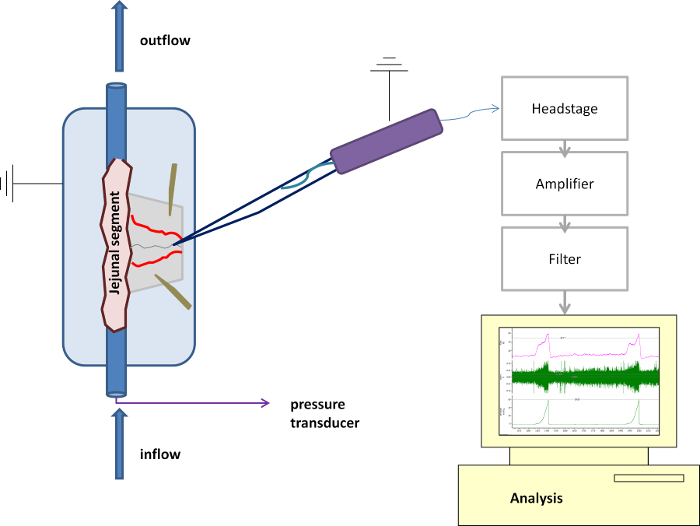
Figure 1: Schematic Overview of the Purpose-built Recording Chamber and Suction Electrode. Detailed overview of the technical setup with the suction electrode and the recording chamber in place. Please click here to view a larger version of this figure.
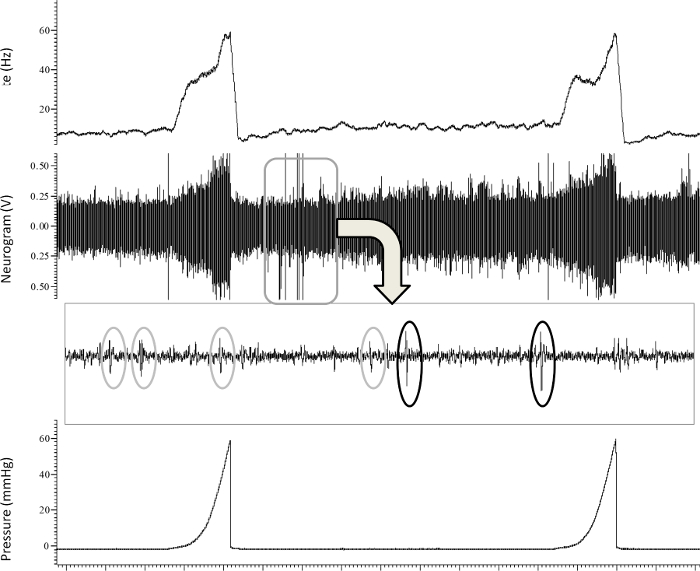
Figure 2: Representative Tracing of the In Vitro Recording of Jejunal Afferent Nerve Activity. Typical recording of jejunal multi-unit afferent nerve activity (imp.sec-1) (upper panel) at baseline and in response to 2 ramp distensions up until 60 mmHg (lower panel), and the subsequent identification (wavemark analysis) of different single-units in the nerve signal (third panel). Please click here to view a larger version of this figure.
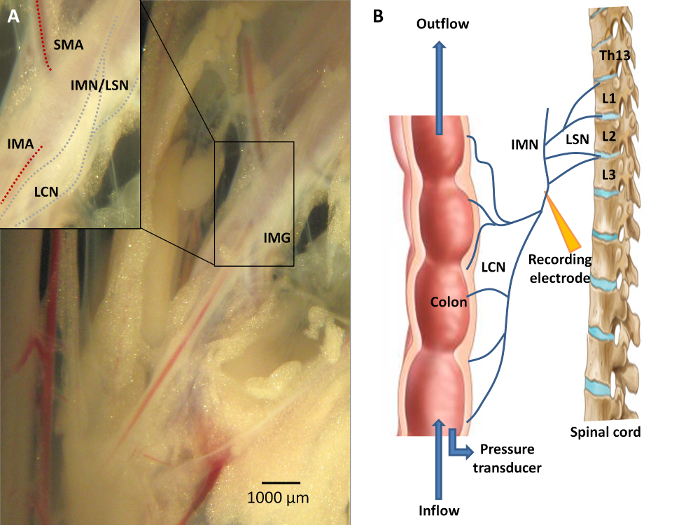
Figure 3: Neuroanatomy of the Colon. A) Sensory information from the colon is conveyed via the lumbar colonic nerves (LCN) towards the central nervous system, with the LCN running in close proximity to the inferior mesenteric artery (IMA). A portion of the fibers from this lumbar colonic nerve will course along the intermesenteric nerve (IMN) to form the lumbar splanchnic nerves (LSN). The inferior mesenteric ganglion (IMG) is located at the origin of the IMA from the abdominal aorta. Recording distally of the IMG is mandatory should researchers wish to study viscerofugal afferent nerve activity. B) A schematic overview of the experimental set-up. Afferent recording of the LCN is performed in an organ both by means of a suction electrode connected to the data acquisition system. Ramp distension can be performed upon closure of the outlet port while continuing the inflow of Krebs solution. Please click here to view a larger version of this figure.
2. Data Acquisition
- Record the nerve activity via a suction electrode connected to a headstage. Amplify (gain 10k) and filter the signal (bandpass 500 – 5,000 Hz) 20.
NOTE: A 50/60 Hz Noise Eliminator should be included in the experimental setup in order to remove 50/60 Hz electrical interference noise signals. The signal is automatically digitized and sampled at 20 kHz for analysis.
3. Analysis5,20
- Report the afferent nerve discharge as the overall number of impulses/second for the entire nerve or use specialized software to perform further analysis, as multi-unit recordings of overall nerve activity contain action potentials of different shape, amplitude and width, corresponding to different nerve units in each afferent fiber (Figure 2).
- Match individual wavemarks to predefined templates, allowing discrimination between single-units. Before allocation of a spike to a certain wave-mark, a signal-to-noise ratio of >2:1 should be enforced.
NOTE: During wavemark analysis, we construct a new template when at least 10 similar spikes are identified by the analysis software. No template is constructed for shapes rarer than 1 in 50 spikes. A spike is matched to a template using principle component analysis when at least 80% of its points are identical to the template spike. Template matching is operator-dependent, and should always be performed by the same researcher in a blinded manner. - Calculate the action potential responses by subtracting the spontaneous activity at baseline (intraluminal pressure of 0 mmHg) from the response during distension at fixed time points (5 mmHg of increment from 0 to 20 mmHg intraluminal pressure, increments of 10 mmHg from 20 mmHg onwards). Measure baseline afferent discharge using a bin size of 10 sec.
- Use the single-unit analysis to classify fibers based upon their discharge profile during ramp distension (Figure 4). In addition, single-unit analysis can be utilized to study the chemosensitive profile of different types of units, as not all types of units will display the same altered firing activity in response to a drug or compound.
- Classify fibers as low threshold fibers ('LT', typically exert increased discharge at the lower distension pressures), high threshold fibers ('HT', exert increased discharge at the highest distension pressures), wide dynamic range fibers ('WDR', demonstrate increased firing during the entire ramp distension) or mechanically insensitive afferent ('MIA', nerve fibers that typically do not respond to ramp distensions) 5;20.
- Express the nerve firing response at 20 mmHg as the percentage of the maximum firing response during distension (LT%) as it reflects the extent of firing at low distension pressure.
NOTE: Therefore, LT fibers are characterized by an LT% > 55%, whereas HT are defined by a value of <15%. Values for WDR units range between 15 and 55% (20). A MIA displays spontaneous afferent firing that is unaffected by distensions.
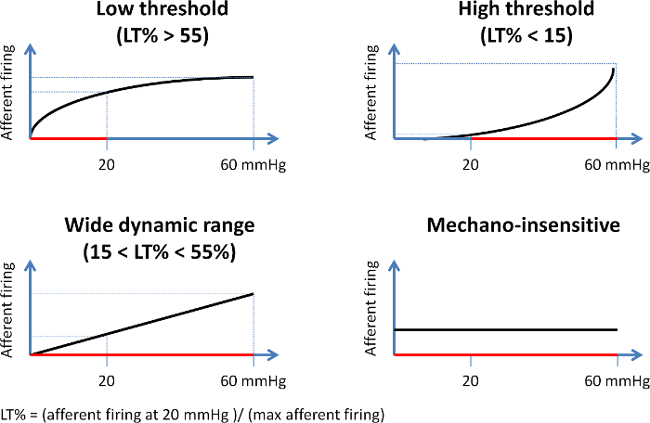
Figure 4: Schematic Representation of the Different Afferent Fiber Units Based on Their Mechanosensitive Profile. Units are classified based upon the percentage (LT%) of their firing rate at 20 mmHg distension pressure compared to the maximum firing response during distension. Low threshold fibers (upper left panel) predominantly display an increased nerve activity at low distension pressures, resulting in an LT% of over 55%. High threshold units (upper right panel) on the contrary only display an increase in firing rate at noxious pressures (%LT < 15). Wide dynamic range fibers (lower left panel) display a gradual increase in nerve activity during the entire distension (%LT ranging between 15 and 55), whereas mechanically insensitive fibers (lower right panel) do not respond to increasing distension pressures. LT%: (afferent firing at 20 mmHg / maximal afferent firing) Please click here to view a larger version of this figure.
Jejunal afferent nerve activity was measured at baseline and in response to ramp distension in 9 eight-week old male OF-1 mice. Animals were housed in groups in standardized conditions (6 animals per cage, 20 – 22 °C, humidity 40 – 50%, 12 hr light-dark cycle) with unlimited access to tap water and regular chow. Jejunal segments of mice displayed irregular spontaneous afferent nerve discharge at baseline at an intraluminal pressure of 0 mmHg (mean spontaneous activity 11.47 ± 3.31 imp/sec).
The jejunal afferent nerve activity increased upon performing ramp distensions up until 60 mmHg. Typically, the increase in afferent nerve activity following the rise in intraluminal pressure is characterized by a biphasic response (Figure 5), consisting of an initial rapid increase in firing activity up until the intraluminal pressure reaches 20 mmHg, which can mainly be attributed to the increased firing rate of low threshold fibers. This is then followed by a plateau phase, after which a second increase in firing activity can be observed from 40 mmHg onwards, representing the activation of predominantly high threshold fibers.
Based upon their waveforms, single-units can be discriminated in each multi-unit recording and classified in one of the aforementioned four categories. In 9 mice, we discriminated 40 different units (4.44 ± 1.01 units/jejunal afferent nerve), with the LT units being the most prevalent ones, followed by WDR and HT fibers (Figure 6). The firing activity of the different units in response to ramp distension can be observed in Figure 7.
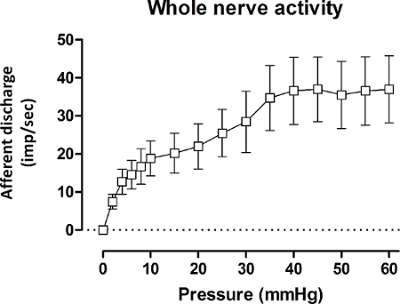
Figure 5: Mesenteric Afferent Nerve Discharge (imp.sec–1) in Wild-type Mice during Ramp Distension. Mesenteric multi-unit afferent nerve discharge (imp/sec-1) in wild-type mice during ramp distension for the whole nerve. Values represent mean afferent discharge ± s.e.m., n = 9 mice. imp.sec-1: impulses per second. Please click here to view a larger version of this figure.
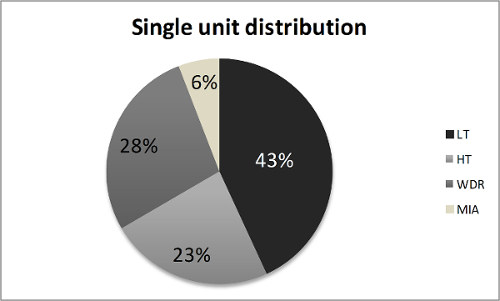
Figure 6: Single Unit Distribution of 40 Units Identified in Jejunal Afferent Nerves from 9 Wild-type Mice. HT: high threshold fiber, LT: low threshold fiber, MIA: mechanically insensitive fiber, WDR: wide dynamic range fiber. Please click here to view a larger version of this figure.
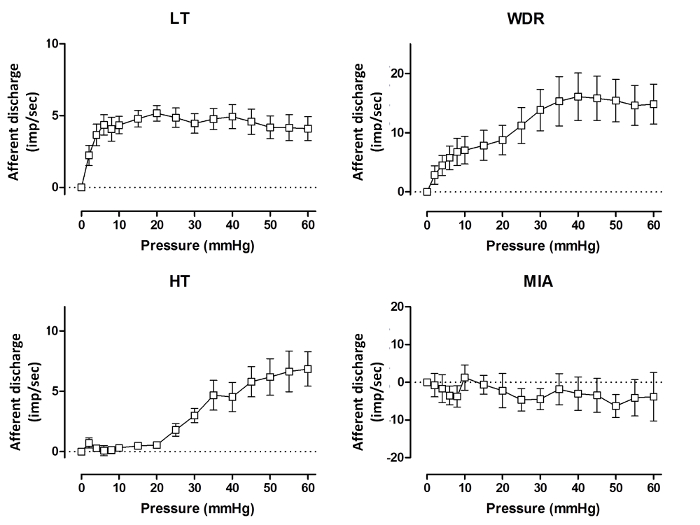
Figure 7: Pressure-response Curves for the Different Types of Subunits in Wild-type Mice. The single-unit afferent nerve discharge (imp.sec-1) from the four different units that can be identified, in wild-type mice during ramp distensions. A low threshold fiber (LT, upper left figure) is characterized by an initial rapid increase in firing activity during distensions, whilst the high threshold fibers (HT, lower left figure) only display increased firing during noxious intraluminal pressures. Wide dynamic range fibers (WDR, upper right figure) show a steady increase in firing activity during the entire distension, and mechano-insensitive afferent fibers (MIA, lower right figure) do not respond to increasing intraluminal pressures. Values represent mean afferent discharge ± s.e.m. imp.sec-1: impulses per second. Please click here to view a larger version of this figure.
