Celler indeholder et væld af biomakromolekyler der konstant interagerer med hinanden. Denne forening giver anledning til komplekser, der deltager i cellulære veje, der er ansvarlige for deres funktion i signaltransduktion, regulering af genekspression og cellevandring blandt andre. Alle protein-protein interaktioner, der forekommer i en celle omfatter et netværk kendt som interactome. I Saccharomyces cerevisiae mere end 70% af dets proteiner er blevet vist at have interagerende partnere 1. Forståelse af interactome af en celle og deres funktioner give relevante oplysninger om den kompleksitet og mangfoldighed af levende organismer. Adskillige metoder er blevet beskrevet til at identificere og karakterisere protein-protein interaktioner. Anderledes højkapacitetsscreening metoder såsom gær-to-hybrid 2, protein-fragment komplementerings- assays 3, affinitetsoprensning 4 koblet til massespektrometri og protein microarrays anvendes til at identificere en interaktion 5,6. Når identificeret, er det nødvendigt at validere det og kan variere fra sag til sag. Typisk er disse eksperimenter involverer forstyrre interaktionen selv på niveau med de enkelte medlemmer af samspillet parret, fx ved gen-sletning eller overekspression af et af proteinerne, og derefter på udkig efter ændringer i egenskaber eller funktion af det andet medlem ved celleniveau. Efterfølgende biofysiske teknikker 7 anvendes til at karakterisere protein-protein-interaktion på det molekylære niveau. Til dette formål er strukturen af proteinkomplekser bestemt ved røntgenkrystallografi, kernemagnetisk resonans og cryo-elektronmikroskopi mens kalorimetri og fluorescensspektroskopi anvendes til kvantitativt og mekanistisk beskrive dem.
I dette arbejde blev fluorescensanisotropi anvendt som en teknik til at karakterisere vekselvirkningen mellem GTPase EFL1 og SBDS-protein. Disse proteiner deltager i syntesen af ribosomer ved at fremme frigivelsen af eukaryot initieringsfaktor 6 fra overfladen af 60S ribosomale subunit 8. Den SBD'er proteinet er muteret i en sygdom kendt som Shwachman-Diamond Syndrome 9 og virker som en guanin nukleotid udveksling faktor for EFL1 faldende dets affinitet for Guanosindifosfat 10,11. Sygdomstilstande mutationer i SBD'er afskaffe interaktion med EFL1 og dermed forhindre dens aktivering.
Fluorescensanisotropi er almindeligt anvendt i biologiske anvendelser til at studere protein-peptid eller protein-nukleinsyre-interaktioner. Den er baseret på princippet om, at en fluorofor ophidset med polariserede lys resulterer i en delvist polariseret emission. Fluorescensanisotropi er defineret ved ligning 1:
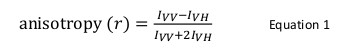
hvor jeg VV og jeg VH er denfluorescensintensiteter af den vertikalt (VV) og horisontalt (VH) polariseret emission, når prøven er spændt med lodret polariseret lys 12. Fluorescensanisotropi er følsom over for faktorer, der påvirker hastigheden af den roterende diffusion af fluoroforen og således afhænger af temperaturen, viskositeten af opløsningen og den tilsyneladende molekylstørrelse af fluoroforen. Den tilsyneladende størrelse af et protein indeholdende en fluorofor øges, når den interagerer med et andet protein og en sådan ændring kan derefter evalueres som en ændring i anisotropi. Mere specifikt vil en fluorofor, der roterer langsomt i opløsning i forhold til dets fluorescerende levetid har et stort I VV værdi og lille I VH-værdi, og derfor vil udvise en relativt stor anisotropi. For fluoroforer der tumler hurtigt i forhold til deres fluorescerende levetid, jeg VV og jeg VH vil være ens og deres anisotropi værdi vil være små 12 </sup> (Figur 1). Endvidere for en god anisotropi signal-støj måling, er det nødvendigt at have en fluorofor med en fluorescenslevetid ligner rotationskorrelationstiden af molekylet af interesse. Ellers er det ikke muligt præcist at registrere forskellen i anisotropi mellem den frie protein og at i komplekset. F.eks anisotropien af en fluorescerende probe med en levetid tæt på 4 ns såsom fluorescein eller rhodamin bundet til en forbindelse med lav molekylvægt på 100 Da er 0,05. Binding til et molekyle på 160 kDa vil øge sin anisotropi værdi til 0,29; en forskel, der kan måles nøjagtigt. I modsætning hertil vil den samme fluorescerende probe involveret i en bindingsreaktion hvis forøgelse molekylstørrelse varierer fra 65 til 1.000 kDa kun resultere i en anisotropi ændring på 0,28 til 0,3, som er for lille til at blive målt nøjagtigt. I dette scenario vil en sonde med en levetid på 400 ns være mere passende 12.
<pclass = "jove_content">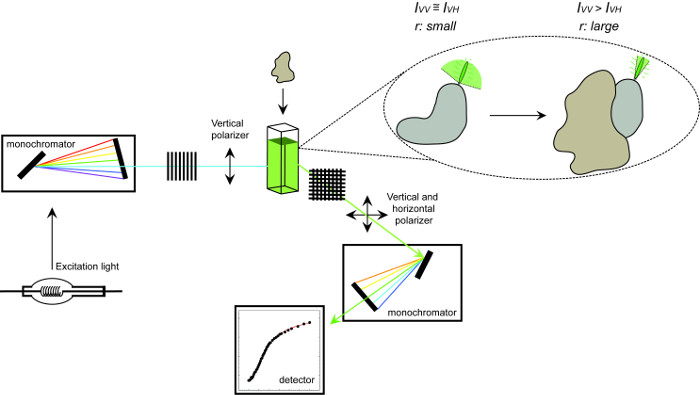
Figur 1. Skematisk gengivelse af det udstyr, der anvendes til at måle fluorescens-anisotropi og proceduren. Skematisk afbildning af udstyr, der anvendes til at udføre et protein-protein-interaktion eksperiment måling af fluorescens-anisotropi. Fluoroforer der tumler hurtigt display lille anisotropi, der øger efter binding til en interaktion partner. Klik her for at se en større version af dette tal.
Fluorescens applikationer kræver tilstedeværelsen af en fluorofor i nogen af de undersøgte molekyler. For at undersøge protein-protein interaktioner er der tre typer af fluoroforer: 1) de tryptophanrester stede i proteinerne, 2) kemisk forbundet fluoroforer og 3) fluorescerende fusionspartnere såsom grønt fluorescerende protein (GFP) og dens derivativer. De fleste proteiner har tryptophanrester af strukturen, således den nemmeste måde at måle interaktion ved at overvåge ændringer i den tilsvarende fluorescens spektre eller ved at overvåge ændringer i fluorescensintensitet af de tryptophanrester. Dog kan tryptophanrester være til stede i begge proteiner komplicerer analysen. På den anden side, for en fluorofor at ændre sine fluorescerende egenskaber skyldes interaktion det skal være placeret på eller nær bindingsstedet og det kunne ændre på selve interaktion. Dette kræver særlig opmærksomhed ved brug af voluminøse fluoroforer såsom GFP. Hvis der kan anvendes ingen af disse fluorophorer til bindingsundersøgelser er det nødvendigt, da, at indføre ekstrinsiske fluoroforer til en af de involverede proteiner. Der findes mange kemisk syntetiserede fluorophorer og kan være kovalent bundet til proteiner via deres reaktive grupper såsom aminogrupper (sidekæde af lysiner eller N-terminus) og thiolgrupperne i cystein. Fluorophore derivater med isothiocyanatgrupperne og succinimidylestere reagerer med amidgrupper mens iodacetamid og maleimid er thiol-reaktive grupper 13. De mest almindelige farvestoffer, der anvendes i fluorescens applikationer er derivater af fluorescein og rhodamin grønne farvestoffer, coumariner, BODIPY fluoroforer og Alexa Fluor farvestoffer. En detaljeret liste over kommercielt tilgængelige fluorophorer og deres anvendelse kan findes i referencer 14,15. For vellykket mærkning, skal den reaktive gruppe eksponeres på overfladen af proteinet, men på grund af det store antal reaktive funktionelle grupper typisk er til stede i polypeptider er det meget svært at få stedspecifik modifikation. Proteinet af interesse i denne undersøgelse, SBD'er, indeholder 5 gratis cysteiner og 33 lysiner der kan resultere i flere mærkning site. Ikke-ensartet mærkning kan påvirke bindende og vil komplicere dataanalyse som forskellige fluoroforen molekyler kan fremkalde forskellige fluorescerende intensitet signaler efter binding. Overcome dette problem, brugte vi blitzen fluoroforen, 4 ', 5'-bis (1,3,2 dithioarsolan-2-yl) fluorescein til site-direkte etiketten SBD'er protein. Dette er en arsenoxide farvestof med en høj affinitet for fire adskilte cysteiner i et motiv kender som Flash-tag bestående af sekvensen CCXXCC hvor X er andet end cystein 16,17 aminosyre. Denne tetracysteine motiv tilsættes til N- eller C-terminalen af proteinet ved gensplejsning sammen med en passende linker til at forhindre forstyrrelse af den samlede foldning af proteinet. Parret bestående af Flash farvestof og Flash-tag blev oprindeligt designet til stedspecifikke label proteiner i levende celler 17, men det kan også bruges til at mærke oprensede proteiner in vitro som det er eksemplificeret her. Yderligere er enzymatiske strategier også blevet udviklet for at muliggøre stedspecifik funktionalisering af proteiner 18.
I dette manuskript beskriver vi nytten af fluorescensanisotropi asa værktøj til at studere protein-protein-interaktioner. Binding kan vurderes ved simpel inspektion af bindende kurveform mens kan fås kvantitative oplysninger fra pasningen af eksperimentelle data.
