תאים מכילים שפע של Biomacromolecules המגיבה כל זמן אחד עם השני. קשר זה מעורר מתחמי כי להשתתף מסלולים הסלולר אחראי לתפקוד שלהם הולכת אותות, בקרת גנים ואת נדידת תאים בין היתר. כל האינטראקציות בין החלבונים המתרחשות בתא יוצרי רשת המכונית interactome. בשנת שמר האפייה יותר מ -70% מחלבוני הוכחו יש אינטראקציה 1 שותפים. הבנת interactome של התא ואת תפקידיהם לספק מידע רלוונטי על המורכבות והמגוון של יצורים חיים. מתודולוגיות מספר תוארו לזהות ולאפיין אינטראקציות חלבון-חלבון. שונות גבוהה באמצעות שיטות לשים כגון שני היברידית שמרים 2, מבחני השלמה-מקטעי חלבון 3, טיהור זיקה 4 מצמידים ספקטרומטריית מסה microarra חלבוןys משמש לזהות אינטראקציה 5,6. לאחר הזיהוי, יש צורך לאמת את זה עשוי להשתנות על בסיס כל מקרה לגופו. בדרך כלל, ניסויים אלה כרוכים לשבש את האינטראקציה עצם ברמה של החברים הבודדים של זוג האינטראקציה, למשל, על ידי מחיקת גן או ביטוי יתר של אחד החלבונים, ולאחר מכן מחפשים שינויים במאפיינים או פונקציה של חבר האחר בבית ברמה התאית. בהמשך לכך, טכניקות biophysical 7 משמשות לאפיין את האינטראקציה בין החלבונים ברמה המולקולרית. לשם כך, המבנה של קומפלקסים חלבונים נקבעים על ידי קריסטלוגרפיה באמצעות קרני רנטגן, תהודה מגנטית גרעינית במיקרוסקופ אלקטרונים cryo- בעוד calorimetry וספקטרוסקופיה הקרינה משמשים כמותית מכניסטי לתאר אותם.
בעבודה זו, אנאיזוטרופיה הקרינה שמשה כטכניקה לאפיין את האינטראקציה בין EFL1 GTPase ואת SBDחלבון S. חלבונים אלה משתתפים בבניית הסינתזה של הריבוזומים באמצעות קידום שחרורו של גורם ייזום איקריוטיים 6 מפני השטח של 60S 8 למקטע ריבוזומלי. החלבון SBDS עובר מוטציה במחלה המכונה 9 תסמונת שווקמן-יהלום ומעשי כגורם חילופי נוקלאוטיד גואנין עבור EFL1 יורדת הזיקה שלה עבור דיפוספט guanosine 10,11. מוטציות Disease in SBDS לבטל את האינטראקציה עם EFL1 ובכך למנוע הפעלה שלה.
אנאיזוטרופיה הקרינה הוא נפוץ יישומים ביולוגיים ללמוד אינטראקציות חלבון-פפטיד או חומצות גרעין-חלבון. היא מבוססת על העיקרון כי fluorophore נרגש עם תוצאות אור מקוטב פליטת מקוטב חלקית. אנאיזוטרופיה הקרינה מוגדרת על ידי משוואה 1:
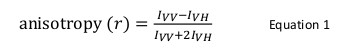
איפה אני VV ואני VH הואעוצמות קרינה של אנכי (VV) ואופקי (VH) מקוטבות פליטה כאשר המדגם הוא נרגש עם מקוטב אנכי אור 12. אנאיזוטרופיה הקרינה היא רגישה לגורמים המשפיעים על השער של דיפוזיה הסיבוב של fluorophore ובכך תלויה בטמפרטורה, את הצמיגות של הפתרון ואת הגודל המולקולרי לכאורה של fluorophore. הגודל לכאורה של חלבון המכיל עליות fluorophore כאשר הוא מקיים אינטראקציה עם חלבון אחר שינוי כזה אז יכול להיות מוערך כשינוי אנאיזוטרופיה. באופן ספציפי יותר, fluorophore שמסתובבת לאט יחסית פתרון לכל חי הניאון שלה תהיה בערך VV אני גדול וערך לי VH קטן ולכן תפגין אנאיזוטרופיה גדול יחסית. עבור fluorophores כי נפילה ביחס במהירות לכל החיים הניאון שלהם, אני VV ואני VH יהיה דומה וערך אנאיזוטרופיה שלהם יהיה קטן 12 </sup > (איור 1). בנוסף, עבור אות אנאיזוטרופיה טובה למדידת רעש, זה הכרחי כדי לקבל fluorophore עם חי שלמי קרינה דומים בפעם מתאם הסיבוב של המולקולה של עניין. אחרת, לא ניתן לרשום את ההבדל במדויק אנאיזוטרופיה בין חלבון חינם וכי במתחם. לדוגמא, אנאיזוטרופיה של בדיקת ניאון עם חיים שלמים קרובים ל -4 NSEC כגון והעמסה או rhodamine מצורפות תרכובת משקל מולקולרית נמוכה של 100 Da היא 0.05. כריכה למולקולה של 160 kDa יגדל ערך אנאיזוטרופיה שלה ל -0.29; הבדל כי ניתן למדוד באופן מדויק. לעומת זאת, באותה הבדיקה הניאון מעורבת תגובה מחייבת גידול אשר בגודל מולקולרי משתנה מ -65 ל 1,000 kDa תגרום רק שינוי אנאיזוטרופיה של 0.28 עד 0.3, שהוא קטן מכדי שיהיה אפשר למדוד במדויק. בתרחיש זה, בדיקה עם חיים שלמים של 400 NSEC תתאים יותר 12.
<p class = "jove_content">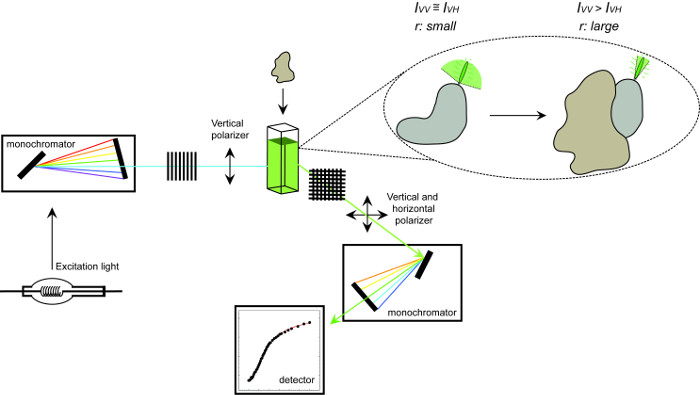
באיור 1. ייצוג סכמטי של ציוד המשמש למדידת אנאיזוטרופיה הקרינה ואת ההליך. ייצוג סכמטי של הציוד המשמש לביצוע אנאיזוטרופיה הקרינה מדידה אינטראקציה בין חלבונים הניסוי. Fluorophores כי נפילת אנאיזוטרופיה קטנה תצוגה מהר המגבירה על קשירת שותף אינטראקציה. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
יישומי קרינה דורשים נוכחות של fluorophore בכל המולקולות למדו. כדי לחקור אינטראקציות חלבון-חלבון ישנם שלושה סוגי fluorophores: 1) שאריות טריפטופן נוכח החלבונים, 2) fluorophores מצורף כימי 3) שותפי היתוך ניאון כגון חלבון פלואורסצנטי ירוק (GFP) ו deriva שלהעזרי למידה. לרובם יש חלבוני שאריות טריפטופן על מבנו, ולכן הדרך הקלה למדוד אינטראקציה היא על ידי מעקב אחר השינויים בספקטרום הקרינה המקביל או על ידי ניטור שינויים בעוצמת הקרינה של שאריות טריפטופן. עם זאת, שאריות טריפטופן עשויות להיות נוכחות בשני החלבונים מסבכים את הניתוח. מצד שני, עבור fluorophore לשינוי תכונות ניאון שלה עקב אינטראקציה זה צריך להיות ממוקם על או ליד אתר הקישור וזה יכול להפריע לאינטראקציה עצמה. זה צריך תשומת לב מיוחדת בעת שימוש fluorophores המגושם כגון GFP. אם אף אחת fluorophores אלה יכולים לשמש למחקרים מחייב יש צורך, אם כן, להציג את fluorophores חיצונית לזו של חלבונים המעורבים. fluorophores רב מסונתזים כימי קיים והוא יכול להיות מחובר קוולנטית חלבונים באמצעות הקבוצות תגובתי שלהם כגון קבוצות האמינות (שרשרת צד של lysines או N- סופי) ואת קבוצות תיאול ב ציסטאין. Fנגזר luorophore עם אסטרים isothiocyanate ו succinimidyl להגיב עם קבוצות אמידות בעוד iodoacetamide ו maleimide קבוצות תיאול-reactive 13. הצבעים הנפוצים ביותר בשימוש ביישומי קרינה הם נגזרים של וההעמסה ואת צבע ירוק rhodamine, coumarins, fluorophores BODIPY וצבעי אלקסה פלואוריד. רשימה מפורטת של fluorophores זמינים מסחרית והשימוש בהם ניתן למצוא אזכור 14,15. עבור תיוג מוצלח, הקבוצה תגובתי חייבת להיחשף על פני השטח של החלבון, אך בשל המספר הגדול של קבוצות פונקציונליות תגובתי בדרך כלל נוכח פוליפפטידים קשה מאוד להשיג שינוי באתר ספציפי. החלבון של עניין במחקר זה, SBDS, מכיל 5 cysteines חינם ו -33 lysines שעלולים לגרום תיוג אתרים מרובים. תיוג לא אחיד עשוי להשפיע על המחייב יסבך ניתוח נתונים כמולקולות fluorophore שונים עשויות לעורר אותות בעצמת ניאון שונים על כריכה. כדי overcome בעיה זו, השתמשנו fluorophore פלאש, 4 ', 5'-bis (1,3,2 dithioarsolan-2-י.ל.) והעמסת לתווית באתר-ישיר החלבון SBDS. זהו צבע arsenoxide עם זיקה גבוהה במשך ארבע cysteines במרווחים מוטיב יודע כמו פלאש-תג המורכב של רצף CCXXCC כאשר X הוא כל חומצת אמינו מלבד ציסטאין 16,17. מוטיב tetracysteine זה מתווסף N- או C- הסופי של החלבון על ידי הנדסה גנטית יחד עם מקשר נאות כדי למנוע את השיבוש של הקפל הכללי של החלבון. הצמד המורכב לצבוע Flash ו- Flash תג תוכנן במקור לחלבוני תווית ספציפי לאתר בתאים חי 17 אבל זה יכול לשמש גם כדי לתייג חלבונים מטוהרים במבחנה כפי שהוא בא לידי ביטוי כאן. בנוסף, אסטרטגיות האנזימטית גם פותחו כדי לאפשר functionalization ספציפית לאתר של חלבונים 18.
בכתב היד הזה אנו מתארים את השימושיות של אנאיזוטרופיה קרינהכלי sa ללמוד אינטראקציות בין חלבונים. כריכה ניתן להעריך על ידי בדיקה פשוטה של הצורה העקומה מחייבת בעוד ניתן להשיג מידע כמוני מן ההתאמה של נתוני הניסוי.
