Le cellule contengono una moltitudine di biomacromolecole che interagiscono costantemente con l'altro. Questa associazione dà luogo a complessi che partecipano alle vie cellulari responsabili per il loro funzionamento nella trasduzione del segnale, regolazione dell'espressione genica e la migrazione delle cellule, tra gli altri. Tutte le interazioni proteina-proteina che si verificano in una cella comprendono una rete nota come dell'interattoma. In Saccharomyces cerevisiae hanno dimostrato più del 70% delle proteine di avere partner interagenti 1. Comprendere la interattoma di una cella e le loro funzioni di fornire informazioni pertinenti sulla complessità e la diversità degli organismi viventi. Diverse metodologie sono state descritte per identificare e caratterizzare le interazioni proteina-proteina. Diverso alto attraverso metodi messi come il lievito doppio ibrido 2, saggi di complementazione proteine frammento 3, affinità di purificazione 4 accoppiata alla spettrometria di massa e microarra proteineys vengono utilizzati per identificare una interazione 5,6. Una volta identificato, è necessario convalidare e puo variare caso per caso. Tipicamente, questi esperimenti comportano interrompere stessa interazione a livello dei singoli membri della coppia interazione, ad esempio, dalla delezione gene o sovraespressione di una delle proteine, e quindi cerca di cambiamenti nelle proprietà o funzione di un altro membro del livello cellulare. Successivamente, tecniche biofisiche 7 sono utilizzati per caratterizzare l'interazione proteina-proteina a livello molecolare. A tal fine, la struttura di complessi proteici sono determinati mediante cristallografia a raggi X, risonanza magnetica nucleare e la microscopia crioelettronica mentre calorimetria e spettroscopia di fluorescenza sono utilizzati per descriverli quantitativamente e meccanicamente.
In questo lavoro, anisotropia di fluorescenza è stato utilizzato come tecnica per caratterizzare l'interazione tra la GTPasi EFL1 e SBDS proteine. Queste proteine partecipano nella sintesi dei ribosomi promuovendo il rilascio di fattore di inizio eucariotico 6 dalla superficie 60S ribosomiale subunità 8. La proteina SBDS è mutato in una malattia nota come Shwachman-Diamond Sindrome 9 e agisce come un fattore di scambio guanina nucleotide per EFL1 diminuendo la sua affinità per guanosindifosfato 10,11. mutazioni malattia in SBDS abolire l'interazione con EFL1 e quindi impediscono la sua attivazione.
Fluorescenza anisotropia è comunemente usato in applicazioni biologiche per studiare la proteina-peptide o interazioni degli acidi nucleici proteine. Si basa sul principio che un fluoroforo eccitato con risultati luce polarizzata in una emissione parzialmente polarizzata. Anisotropia di fluorescenza è definito dall'equazione 1:
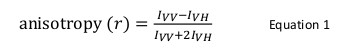
dove ho vv e mi VH sono ilintensità di fluorescenza della verticale (VV) e in orizzontale (VH) polarizzati emissione quando il campione è eccitato con luce polarizzata verticalmente 12. Anisotropia di fluorescenza è sensibile a fattori che influenzano la velocità di diffusione rotazionale del fluoroforo e dipende dalla temperatura, la viscosità della soluzione e la dimensione molecolare apparente del fluoroforo così. La dimensione apparente di una proteina contenente un fluoroforo aumenta quando interagisce con un'altra proteina e tale modifica può essere valutato come un cambiamento di anisotropia. Più specificamente, un fluoroforo che ruota lentamente in soluzione rispetto alla sua durata fluorescente avrà un valore I VV e piccolo valore I VH e pertanto presentano una relativamente grande anisotropia. Per fluorofori che scendono rapidamente rispetto al loro ciclo di vita fluorescente, ho Vv e VH saranno simili e il loro valore anisotropia sarà piccolo 12 </sup> (Figura 1). Inoltre, per un buon segnale anisotropia di misura del rumore, è necessario avere un fluoroforo con una vita di fluorescenza simile al tempo di correlazione rotazionale della molecola di interesse. Altrimenti, non è possibile registrare accuratamente la differenza di anisotropia tra la proteina libero e che nel complesso. Ad esempio, l'anisotropia di una sonda fluorescente con una durata quasi 4 nsec come fluoresceina o rodamina collegati a un composto a basso peso molecolare di 100 Da è 0.05. Associazione a una molecola di 160 kDa aumenterà il suo valore anisotropia a 0,29; una differenza che può essere misurata con precisione. Al contrario, la stessa sonda fluorescente coinvolto in una reazione di legame cui incremento di peso molecolare varia da 65 a 1000 kDa comporterà solo un cambiamento anisotropia di 0,28 e 0,3, che è troppo piccola per essere misurata con precisione. In questo scenario, una sonda con una durata di 400 nanosecondi sarebbe più adatto 12.
<pclass = "jove_content">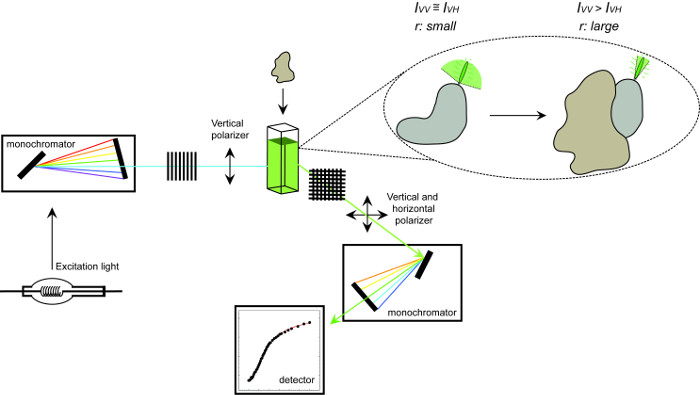
Figura 1. Rappresentazione schematica delle apparecchiature utilizzate per misurare anisotropia di fluorescenza e la procedura. Rappresentazione schematica dell'attrezzatura utilizzata per eseguire una anisotropia di fluorescenza misurazione proteina-proteina interazione esperimento. Fluorofori che scendono visualizzazione rapida piccolo anisotropia che aumenta dopo il legame a un partner di interazione. Cliccate qui per vedere una versione più grande di questa figura.
applicazioni fluorescenza richiedono la presenza di un fluoroforo in una qualsiasi delle molecole studiate. Per studiare le interazioni proteina-proteina ci sono tre tipi di fluorofori: 1) i residui di triptofano presenti nelle proteine, 2) fluorofori allegate chimicamente e 3) partner di fusione fluorescenti come la proteina fluorescente verde (GFP) e la sua Derivative. La maggior parte delle proteine hanno residui di triptofano sulla sua struttura, così il modo più semplice per misurare un'interazione è monitorando i cambiamenti nel corrispondente spettri di fluorescenza o monitorando i cambiamenti nell'intensità di fluorescenza dei residui di triptofano. Tuttavia, residui di triptofano possono essere presenti in entrambe le proteine complicare l'analisi. D'altra parte, per un fluoroforo per modificare le proprietà fluorescenti dovuto all'interazione deve essere situato sopra o vicino al sito di legame e potrebbe interferire con l'interazione stessa. Questo richiede particolare attenzione quando si utilizza fluorofori ingombranti come GFP. Se nessuno di questi fluorofori può essere utilizzato per studi di legame è necessario, quindi, di introdurre fluorofori estrinseci a una delle proteine coinvolte. Molti fluorofori sintetizzati chimicamente esistono e possono essere covalentemente attaccati alle proteine attraverso i loro gruppi reattivi come i gruppi amminici (catena laterale di lisine o N-terminale) ei gruppi tiolici in cisteina. Fderivati luorophore con isotiocianato e succinimidyl esteri reagiscono con gruppi ammidici mentre Iodoacetamide e maleimmide sono gruppi tiolo-reattivi 13. I coloranti più comuni utilizzati in applicazioni in fluorescenza sono derivati del fluoresceina ei coloranti verdi rodamina, cumarine, fluorofori BODIPY e coloranti Alexa Fluor. Un elenco dettagliato dei fluorofori disponibili in commercio e il loro uso può essere trovato in riferimenti 14,15. Per l'etichettatura di successo, il gruppo reattivo deve essere esposto sulla superficie della proteina, ma a causa del gran numero di gruppi funzionali reattivi tipicamente presenti in polipeptidi è molto difficile ottenere modificazioni sito-specifica. La proteina di interesse in questo studio, SBDS, contiene 5 cisteine libere e 33 lisine che possono provocare l'etichettatura più siti. l'etichettatura non uniforme può influenzare il legame e complicherà l'analisi dei dati come diverse molecole fluorofori possono suscitare diversi segnali di intensità fluorescente su vincolante. a Overcome questo problema, abbiamo usato il fluoroforo flash, 4 ', 5'-bis (1,3,2 dithioarsolan-2-il) fluoresceina all'etichetta sito-diretta la proteina SBDS. Questo è un colorante arsenoxide con alta affinità per quattro cisteine distanziati in un motivo conosciuto come FlAsH-tag costituito dalla CCXXCC sequenza in cui X è un qualsiasi amminoacido diverso cisteina 16,17. Questo motivo tetracysteine viene aggiunto N- o C-terminale della proteina mediante ingegneria genetica insieme ad un linker opportuno evitare l'interruzione della piega complessiva della proteina. La coppia che consiste di Flash tintura e Flash-tag è stato originariamente progettato per le proteine di etichette site-specific nelle cellule viventi 17, ma può anche essere usato per etichettare le proteine purificate in vitro come è esemplificato qui. Inoltre, sono stati sviluppati strategie enzimatici per consentire funzionalizzazione site-specific di proteine 18.
In questo manoscritto descriviamo l'utilità di anisotropia di fluorescenza unTool SA per studiare le interazioni proteina-proteina. Binding può essere valutata semplice ispezione della forma vincolante curva mentre le informazioni quantitative può essere ottenuta dalla misura dei dati sperimentali.
