I 1998 Fire og Mello rapporterede, at dobbeltstrenget RNA (dsRNA) kan fremkalde hæmning af gen funktion i Caenorhabditis elegans 1. Denne reaktion udløst af dsRNA blev opkaldt RNA-interferens (RNAi), og sådan RNAi-medieret gen silencing blev rapporteret at være bevaret i dyr, planter og svampe 2-7. I planter og nogle dyr, RNAi fungerer systemisk, hvilket betyder, at virkningen kan sprede sig til andre celler / væv, hvor dsRNA ikke er direkte indført (gennemgået i 8-10). Forskere har gjort brug af denne endogene cellulære RNAi respons ved at designe dsRNA'er at målrette gener af interesse, og dermed vælte gen funktion uden direkte at manipulere genomet (revideret i 11-14).
RNAi er et stærkt værktøj til funktionelle undersøgelser på grund af følgende fordele. Første, selv med minimal gensekvensinformation, et gen kan målrettes ved hjælp af RNAi. Dette er især vigtigt for studies af ikke-modelorganismer mangler genomiske eller transkriptom data. For det andet, i organismer hvor RNAi respons er robust systemisk, RNAi-medieret gen knockdown kan udføres på næsten alle udviklingstrin. Denne funktion er meget nyttig til undersøgelse af funktionen af pleiotrope gener. For det tredje, i nogle tilfælde spredes RNAi effekter til gonaderne og afkom, således at fænotyper hos afkommet 15,16. Dette fænomen, kendt som forældrenes RNAi (pRNAi), er særligt fordelagtigt for gener påvirker fosterudviklingen, så talrige afkom af en enkelt indsprøjtet forælder kan undersøges uden direkte manipulation af æg. Af disse grunde, pRNAi er den foretrukne fremgangsmåde. Men hvis pRNAi er ineffektiv, for eksempel for gener, der er nødvendige for oogenesen derefter embryonale RNAi (eRNAi) skal anvendes. For det fjerde kan RNAi anvendes til at frembringe hvad der svarer til en allelisk serie ved, at mængden af dsRNA leverede kan varieres over et område for at frembringe svage til stærke defekter. En sådan graduering af fænotyper kan være nyttigt for forståelsen genfunktioner når genet er involveret i en kompleks proces og / eller fuldstændigt tab af funktion er dødelig. Femte, levering af dsRNA er generelt let og muligt, især i dyr, der viser robuste systemiske RNAi svar. dsRNA kan indføres ved mikroinjektion 1,5, fodring / indtagelse 17,18, iblødsætning, 19,20 og virus / bakterie-medieret levering 21,22. Sjette, i modsætning til nogle genmålretning / redigering metoder, er der ingen grund til at screene for organismer, der bærer mutationen eller foretage genetiske krydsninger at generere homozygoter ved brug RNAi. Derfor sammenlignet med mange andre teknikker til at studere genfunktion, RNAi er hurtig, billig, og kan anvendes til store skærme 23-25.
Den brede anvendelighed af RNAi tilvejebringer midler til at udføre funktionelle studier i en lang række organismer, udvide række arter til rådighed til undersøgelse beyond de traditionelle modelsystemer for hvilke der er udviklet genetiske værktøjer. For eksempel er undersøgelser med anvendelse af ikke-modelsystemer forpligtet til at give indblik i udviklingen af gener og gen-netværk ved at sammenligne funktionerne i ortologer fra arter, der repræsenterer forskellige udviklingsprojekter tilstande eller udstilles distinkte morfologiske træk 26-29. Disse typer af undersøgelser vil give en bedre forståelse af den biologiske mangfoldighed, med konsekvenser for både anvendt forskning og grundforskning.
Som den største dyregruppe på planeten, insekter giver en stor mulighed for at udforske de mekanismer, der ligger mangfoldighed. Derudover insekter er generelt små, har korte livscyklus, høj frugtbarhed, og er lette at opdrætte i laboratoriet. I de seneste to årtier har RNAi blevet anvendt med succes i insekter spænder ordrer, herunder Diptera (sande fluer) 5, Lepidoptera (sommerfugle og møl) 30, Coleoptera (biller) 16,31, Hymenoptera (sawfløgne, hvepse, myrer og bier) 32, Hemiptera (sande bugs), Isoptera (termitter) 34, Blattodea (kakerlakker) 35, Orthoptera (fårekyllinger, græshopper, græshopper og katydids) 36 og Phthiraptera (lus) 37. Vellykket anvendelse af RNAi har givet funktionelle data for studier af mønstre i tidlig embryogenese (anterior-posterior akse 32, dorsal-ventral akse 28, segmentering 26,38), kønsbestemmelse 39,40, kitin / neglebånd biosyntese 41, ecdyson signalering 42, social adfærd 43, og mere. RNAi metoder udviklet til forskellige insektarter kan være af yderligere fordel ved, at de sandsynligvis vil være nyttige til skadedyrsbekæmpelse (gennemgået i 44-46). RNAi effekter vil være genspecifikke samt artsspecifik, så længe ikke-konserverede regioner valgt til målretning. For gavnlige insektarter som honningbier og silkeorme, rettet mod gener vitale for overlevelsevira eller parasitter at kontrollere infektion kan tilvejebringe en ny strategi til at beskytte disse arter 47,48.
Dermestes maculatus (D. maculatus), almindelig betegnelse skjul bille, fordeles over hele verden med undtagelse af Antarktis. Som holometabolous insekt, D. maculatus livscyklus omfatter embryonale, larve, puppe og voksne stadier (Figur 1). Fordi det feeds på kødet, er D. maculatus bruges i museer at skeletonize døde dyr og retsmedicinske Entomologer kan bruge det til at vurdere dødstidspunktet 49,50. D. maculatus feeds på animalske produkter, herunder kroppe, tørret kød, ost og den pupper / kokoner af andre insekter og forårsager dermed skade på husholdninger, opbevarede fødevarer, og silke, ost, og kødbranchen 51,52. Anvendelse RNAi i denne bille kunne give en effektiv og miljøvenlig måde at minimere dens økonomiske virkninger. Vores laboratorium har brugt D. maculatus som en ny model insekt at studere segmentering 53. Ud over at være modtagelig for lab opdræt, D. maculatus er af interesse for grundforskning, da det er et mellemprodukt-kim udvikler, hvilket gør det til et nyttigt art at studere overgangen mellem kort og langt kim udvikling.
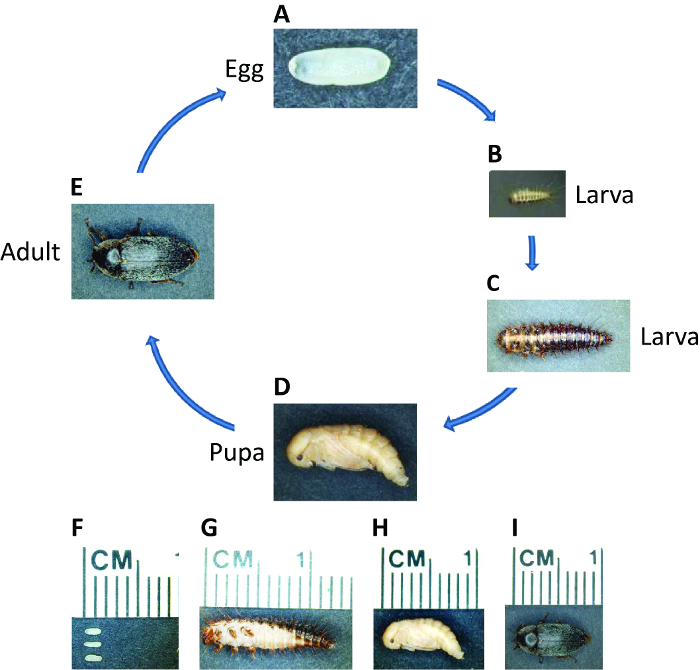
Figur 1: Life Cycle of D. maculatus. Fotografier af D. maculatus på forskellige livsstadier, som angivet. Den livscyklus fra æg til voksen tager tre uger ved 30 ° C, men længere ved lavere temperaturer. (A, F) frisk fastsat embryoner er hvide til lys gul og ovale, ca. 1,5 mm i længde. Embryogenese tager ~ 55 timer ved 30 ° C. (B, C og G) Larver har mørke pigmenterede striber og er dækket med setae. Larver gå gennem flere instars afhængig af miljøet, og deres længde kan strække sig op til over 1 cm. (D, H) </strong> Unge pupper er lys gul. Pupation tager ~ 5 – 7 dage ved 30 ° C. (E, I) Kort efter eclosion synes mørk pigmentering i den voksne bille krop. Voksne kan leve op til flere måneder og en kvindelig kan lægge hundredvis af embryoner i hendes levetid. Klik her for at se en større version af dette tal.
Tidligere viste vi, at RNAi er effektivt til at vælte genfunktion i D. maculatus 53. Her vores erfaring opdræt D. maculatus kolonier i laboratoriet deles sammen med trin-for-trin protokoller for både embryonale og forældreorlov RNAi set-up, injektion, efter injektion pleje, og fænotypisk analyse. De dsRNA-medieret gen Knockdown og analysemetoder indført her ikke kun give detaljerede oplysninger til adressering spørgsmål i D. maculatus, men også have potentiel betydning for anvendelse af RNAi i andre ikke-model bille / insektarter.
