בשנת 1998, אש מלו דיווחו כי RNA פעמיים תקועים (dsRNA) יכול לגרום עיכוב של פונקציה גנים elegans Caenorhabditis 1. תגובה זו מופעלת על ידי dsRNA נקראה התערבות RNA (RNAi), ו להשתקת גנים RNAi בתיווך כזה נמסר ישומרו ב בעלי חיים, צמחים, ופטריות 2-7. בצמחים וכמה חיות, פונקציות RNAi מערכתית, כלומר ההשפעה יכולה להתפשט תאים / רקמות אחרות שבהן dsRNA לא הציג באופן ישיר (הנסקרת ב 8-10). מדענים עשו שימוש בתגובה RNAi הסלולר אנדוגני זאת על ידי תכנון dsRNAs למקד את הגנים של עניין, ובכך דריסה תפקוד הגן ללא מניפולציה של הגנום ישירות (הנסקרת ב 11-14).
RNAi הוא כלי רב עצמה עבור מחקרים תפקודיים בשל היתרונות הבאים. ראשית, אפילו עם מידע רצף הגן מינימלי, גן יכול להיות ממוקד באמצעות RNAi. זה חשוב במיוחד עבור studies של אורגניזמים שאינם מודל חסר הנתונים הגנומי או transcriptomic. שנית, באורגניזמים שבו תגובת RNAi היא מערכתית וחסון, מציאת גן בתיווך RNAi ניתן לבצע כמעט בכל שלב התפתחותי. תכונה זו שימושית מאוד ללימוד הפונקציה של גני pleiotropic. שלישית, במקרים מסוימים, תופעות RNAi להתפשט אל בלוטות המין וצאצאים, כך פנוטיפים הם נצפו בצאצאים 15,16. תופעה זו, המכונית RNAi ההורי (pRNAi), יש יתרון במיוחד עבור גני ההשפיע התפתחות עוברית, כמו צאצאים רבים המיוצרים על ידי הורה מוזרק אחת יכולים להיבחן ללא מניפולציה ישירה של ביצים. מסיבות אלה, pRNAi היא השיטה של בחירה. עם זאת, אם pRNAi אינו יעיל, למשל עבור הגנים הדרושים oogenesis, אז העוברי RNAi (eRNAi) חייב לשמש. הרביעית, RNAi יכול לשמש כדי ליצור את המקבילה של סדרת אללים ב שכמות dsRNA לגידולים יכולות להיות מגוונות על פני טווח לייצר חלש לפגמים חזקים. כזה הדרגתיות של פנוטיפים יכולה להיות מועילה להבנת תפקוד גן כאשר הגן מעורב תהליך מורכב ו / או אובדן מוחלט של פונקציה הוא קטלני. חמישי, מסירת dsRNA קלה בדרך כלל ריאלית, במיוחד בחיות מראות תגובות RNAi מערכתיות חזקות. dsRNA יכול להיות מוצג על ידי microinjection 1,5, האכלה / בליעה 17,18, השריה, 19,20 ו וירוס / מסירה בתיווך חיידקים 21,22. השישית, בניגוד לשיטות מיקוד / עריכה כמה גן, אין צורך למסך אורגניזמים נושא את המוטציה או לבצע צלבים גנטיים לייצר הומוזיגוטים בעת שימוש RNAi. לכן, לעומת טכניקות רבות אחרות לחקר תפקוד גן, RNAi הוא מהיר, זול, והוא יכול להיות מיושם על מסכים גדול בקנה מידה 23-25.
השירות הרחב של RNAi מספק אמצעים לבצע מחקרים פונקציונליים במגוון רחב של אורגניזמים, הרחבת סל מינים לרשותנו בעבודה beyonד מערכות המודל המסורתי עבורו פותחו כלים גנטיים. לדוגמא, מחקרים באמצעות מערכות שאינם מודל נדרשים לתת תובנה על האבולוציה של גני רשתות גנים ידי השוואת הפונקציות של orthologs ממינים המייצגות מצבי פיתוח שונים או מפגין תכונות מורפולוגיות ברורות 26-29. אלו סוגים של מחקרים יספקו הבנה טובה יותר של מגוון ביולוגי, עם שפעות הוא למחקר יישומים ובסיסי.
להיות קבוצת החיה הגדולה ביותר בכדור הארץ, חרקים מספקים הזדמנות מצוינת לחקור את המנגנונים מגוונים בסיסית. בנוסף, החרקים קטנים בדרך כלל, יש מחזור חיים קצר, פוריות גבוהה, והם קלים אחוריים במעבדה. בשני העשורים האחרונים, RNAi יושם בהצלחה חרקים פורש הזמנות, כולל Diptera (זבובים נכון) 5, פרפראים (פרפרים ועשים) 30, Coleoptera (חיפושיות) 16,31, דבוראים (sawfשקרים, צרעות, נמלים ודבורים) 32, פשפשאים (באגים נכון), Isoptera (טרמיטים) 34, Blattodea (ג'וקים) 35, חגבאים (צרצרים, חגבים, ארבה, ו katydids) 36 ו Phthiraptera (כינים) 37. יישום מוצלח של RNAi ספק נתונים תפקודיים ללימודים של דפוסים ב העובר מוקדם (posterior-קדמי ציר 32, ציר-גחון הגבה 28, פילוח 26,38), קביעת הזוויג 39,40, כיטין / לציפורן ביוסינתזה 41, ecdysone איתות 42, התנהגות חברתית 43, ועוד. שיטות RNAi שפותחו עבור מיני חרקים שונים עשויות להועיל נוסף כי הם צפויים להיות שימושיים עבור דברה (הנסקרת ב 44-46). תופעות RNAi תהיינה גן ספציפי כמו גם מינים ספציפיים, כל עוד אזורים שאינם שמורה נבחרים למיקוד. עבור מינים של חרקים מועילים כמו דבורי דבש תולעי משי, מיקוד גנים חיוניים להישרדותו שלוירוסים או טפילים לשלוט הזיהום עלול לספק אסטרטגיה חדשנית להגן על מינים אלה 47,48.
Dermestes maculatus (ד maculatus), חיפושית להסתיר השם הנפוץ, מופץ ברחבי העולם למעט אנטארקטיקה. כמו חרק holometabolous, מחזור חי ד maculatus כולל עוברי, זחל, גלמים, ושלבי מבוגר (איור 1). כי זה נזון בשר, ד maculatus משמש במוזיאונים כדי skeletonize חיות מתות חוקרי חרקים משפטיים יכולים להשתמש בו כדי להעריך את זמן המוות 49,50. ד maculatus נזון מוצרים מן החי כוללים פגרים, בשר מיובש, גבינה, ואת הגלמים / פקעות של חרקים אחרים ובכך גורם לפגיעה במשקי בית, מזון מאוחסן, ואת המשי, גבינה, ותעשיות בשר 51,52. החלת RNAi ב חיפושית זו יכולה לספק דרך יעילה וידידותית לסביבה כדי למזער את ההשפעה הכלכלית שלה. המעבדה שלנו השתמשה ד maculatus כמו מ 'חדשחרק אודל ללמוד פילוח 53. בנוסף להיותו מקובל גידול במעבדה, ד maculatus הוא עניין למחקר בסיסי כמו זה הינה יזמית ביניים-נבט, מה שהופך אותו זן כדאי ללמוד את המעבר בין קצר ופיתוח-נבט ארוך.
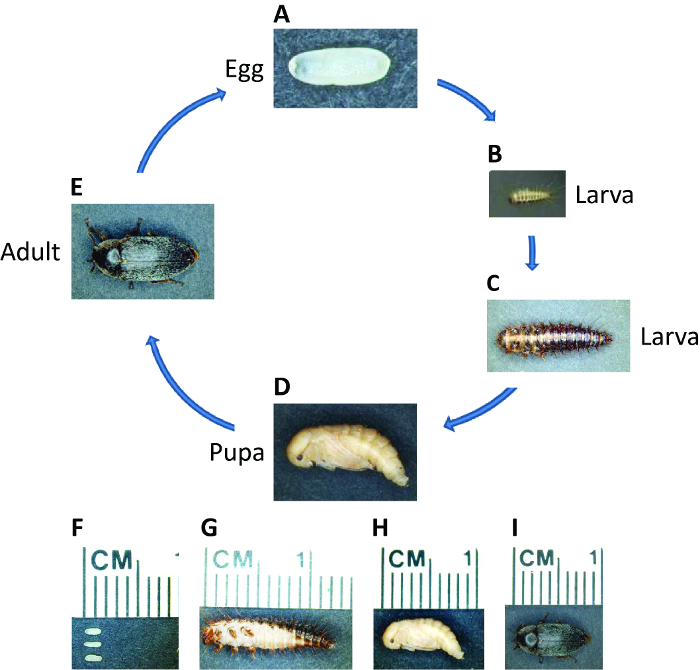
איור 1: מחזור החיים של ד maculatus. תצלומים של ד maculatus בשלבים שונים בחיים, כפי שצוין. מחזור החיים מביצה לבוגר לוקח שלושה שבועות על 30 מעלות צלזיוס אבל כבר בטמפרטורות נמוכות. (A, F) טרי עוברים הניח הם מלבן לצהוב סגלגל בהיר, כ 1.5 מ"מ אורך. העובר לוקח ~ 55 שעות ב 30 מעלות צלזיוס. (B, C ו- G) זחלים יש פסי פיגמנט כהים מכוסים setae. הזחלים לעבור instars מספר בהתאם לסביבה ואורכם יכול להימשך עד מעל 1 ס"מ. (D, H) </strאונג> גלמים צעירים הם צהובים בהירים. ההתגלמות לוקחת ~ 5 – 7 ימים ב 30 מעלות צלזיוס. (E, I) זמן קצר לאחר eclosion, פיגמנטציה כהה מופיעה מעל גוף החיפושית המבוגרת. מבוגרים יכולים לחיות עד מספר חודשים ונקבה אפשר לגולל מאות עוברים במהלך חייה. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
בעבר, הראינו כי RNAi יעיל דריסת תפקוד גן ב ד maculatus 53. הנה הניסיון שלנו לגדל מושבות ד maculatus במעבדה משותף יחד עם צעד-אחר-צעד פרוטוקולים הן עובריים והורית RNAi הגדרת, זריקה, טיפול לאחר ההזרקה, וניתוח פנוטיפי. מציאת גן dsRNA בתיווך ושיטות ניתוח הציגו כאן לא רק לספק מידע מפורט לטיפול שאלות ב ד maculatus, אבל יש גם משמעות פוטנציאל FOr החלת RNAi ב חיפושית שאינם מודל אחר / מיני חרקים.
